EGE เกี่ยวกับชีววิทยางาน C5 การเลือกงานของเซลล์วิทยา องค์ประกอบมาโครและการติดตาม ข้อมูลทางพันธุกรรมในเซลล์ ยีนรหัสพันธุกรรมและคุณสมบัติของมัน ลักษณะของเมทริกซ์ธรรมชาติของปฏิกิริยาการสังเคราะห์การสังเคราะห์ Biosynthesis ของโปรตีนและกรดนิวคลีอิก
D. A. Solovkov ผู้สมัครงานวิทยาศาสตร์ชีวภาพ
งานที่มีให้เลือกนี้มีงาน Cytology ประเภทหลักทั้งหมดที่พบในการใช้งานและมีวัตถุประสงค์หลักเพื่อเตรียมผู้สมัครอย่างอิสระเพื่อแก้ไขภารกิจ C5 ในการสอบ เพื่อความสะดวกงานนี้จัดกลุ่มตามส่วนหลักและหัวข้อที่รวมอยู่ในโปรแกรมชีววิทยา (ส่วน "เซลล์วิทยา") จุดจบแสดงคำตอบสำหรับการทดสอบตัวเอง
ตัวอย่างงานของประเภทแรก
ตัวอย่างของงานประเภทที่สอง
ตัวอย่างของงานประเภทที่สาม
- ชิ้นส่วนของหนึ่งในโซ่ DNA มีโครงสร้างต่อไปนี้: Aagzgtgtzg สร้างบนมันและ rna และกำหนดลำดับของกรดอะมิโนในส่วนของโมเลกุลโปรตีน (สำหรับการใช้งานตารางรหัสพันธุกรรม)
- ส่วนของหนึ่งในโซ่ DNA มีโครงสร้างต่อไปนี้: Catsatsattsggagat สร้างบนมันและ rna และกำหนดลำดับของกรดอะมิโนในส่วนของโมเลกุลโปรตีน (สำหรับการใช้งานตารางรหัสพันธุกรรม)
- ส่วนของหนึ่งในโซ่ DNA มีโครงสร้างต่อไปนี้: Agttttstggcaa สร้างบนมันและ rna และกำหนดลำดับของกรดอะมิโนในส่วนของโมเลกุลโปรตีน (สำหรับการใช้งานตารางรหัสพันธุกรรม)
- ชิ้นส่วนของหนึ่งในโซ่ DNA มีโครงสร้างต่อไปนี้: Gattazthantag สร้างบนมันและ rna และกำหนดลำดับของกรดอะมิโนในส่วนของโมเลกุลโปรตีน (สำหรับการใช้งานตารางรหัสพันธุกรรม)
- ส่วนของหนึ่งในโซ่ DNA มีโครงสร้างต่อไปนี้: ctatcshtctc สร้างบนมันและ rna และกำหนดลำดับของกรดอะมิโนในส่วนของโมเลกุลโปรตีน (สำหรับการใช้งานตารางรหัสพันธุกรรม)
- ส่วนของหนึ่งในโซ่ DNA มีโครงสร้างต่อไปนี้: Aagctazagats สร้างบนมันและ rna และกำหนดลำดับของกรดอะมิโนในส่วนของโมเลกุลโปรตีน (สำหรับการใช้งานตารางรหัสพันธุกรรม)
- ส่วนของหนึ่งในโซ่ DNA มีโครงสร้างต่อไปนี้: ggtgztsgaaag สร้างบนมันและ rna และกำหนดลำดับของกรดอะมิโนในส่วนของโมเลกุลโปรตีน (สำหรับการใช้งานตารางรหัสพันธุกรรม)
- ชิ้นส่วนของหนึ่งในโซ่ DNA มีโครงสร้างต่อไปนี้: Tszzgthaattsg สร้างบนมันและ rna และกำหนดลำดับของกรดอะมิโนในส่วนของโมเลกุลโปรตีน (สำหรับการใช้งานตารางรหัสพันธุกรรม)
ตัวอย่างของภารกิจของประเภทที่สี่
- ชิ้นส่วนและ RNA มีโครงสร้างต่อไปนี้: Gaugaguatsuzaaaa กำหนด Anti-Codons T-RNA และลำดับของกรดอะมิโนที่เข้ารหัสในส่วนนี้ นอกจากนี้เขียนชิ้นส่วนของโมเลกุล DNA ที่สิ่งนี้และ RNA ถูกสังเคราะห์ (สำหรับการใช้งานกับตารางของรหัสพันธุกรรม)
- ชิ้นส่วนและ RNA มีโครงสร้างต่อไปนี้: Tsagguuuzzzurgg กำหนด Anti-Codons T-RNA และลำดับของกรดอะมิโนที่เข้ารหัสในส่วนนี้ นอกจากนี้เขียนชิ้นส่วนของโมเลกุล DNA ที่สิ่งนี้และ RNA ถูกสังเคราะห์ (สำหรับการใช้งานกับตารางของรหัสพันธุกรรม)
- ส่วนและ RNA มีโครงสร้างต่อไปนี้: Uguutsaauaggaagg กำหนด Anti-Codons T-RNA และลำดับของกรดอะมิโนที่เข้ารหัสในส่วนนี้ นอกจากนี้เขียนชิ้นส่วนของโมเลกุล DNA ที่สิ่งนี้และ RNA ถูกสังเคราะห์ (สำหรับการใช้งานกับตารางของรหัสพันธุกรรม)
- ส่วนและ RNA มีโครงสร้างต่อไปนี้: tzcczaatsazgzz กำหนด Anti-Codons T-RNA และลำดับของกรดอะมิโนที่เข้ารหัสในส่วนนี้ นอกจากนี้เขียนชิ้นส่วนของโมเลกุล DNA ที่สิ่งนี้และ RNA ถูกสังเคราะห์ (สำหรับการใช้งานกับตารางของรหัสพันธุกรรม)
- ชิ้นส่วนและ RNA มีโครงสร้างต่อไปนี้: Aqaguggzzzazzz กำหนด Anti-Codons T-RNA และลำดับของกรดอะมิโนที่เข้ารหัสในส่วนนี้ นอกจากนี้เขียนชิ้นส่วนของโมเลกุล DNA ที่สิ่งนี้และ RNA ถูกสังเคราะห์ (สำหรับการใช้งานกับตารางของรหัสพันธุกรรม)
- ชิ้นส่วนและ RNA มีโครงสร้างต่อไปนี้: Gatzagatsutziagutsu กำหนด Anti-Codons T-RNA และลำดับของกรดอะมิโนที่เข้ารหัสในส่วนนี้ นอกจากนี้เขียนชิ้นส่วนของโมเลกุล DNA ที่สิ่งนี้และ RNA ถูกสังเคราะห์ (สำหรับการใช้งานกับตารางของรหัสพันธุกรรม)
- ชิ้นส่วนและ RNA มีโครงสร้างต่อไปนี้: Austzugaacentsgua กำหนด Anti-Codons T-RNA และลำดับของกรดอะมิโนที่เข้ารหัสในส่วนนี้ นอกจากนี้เขียนชิ้นส่วนของโมเลกุล DNA ที่สิ่งนี้และ RNA ถูกสังเคราะห์ (สำหรับการใช้งานกับตารางของรหัสพันธุกรรม)
- ชิ้นส่วนและ RNA มีโครงสร้างต่อไปนี้: gccaggzcauuuuu กำหนด Anti-Codons T-RNA และลำดับของกรดอะมิโนที่เข้ารหัสในส่วนนี้ นอกจากนี้เขียนชิ้นส่วนของโมเลกุล DNA ที่สิ่งนี้และ RNA ถูกสังเคราะห์ (สำหรับการใช้งานกับตารางของรหัสพันธุกรรม)
- ชิ้นส่วนและ RNA มีโครงสร้างต่อไปนี้: Gutuauguatsuuats กำหนด Anti-Codons T-RNA และลำดับของกรดอะมิโนที่เข้ารหัสในส่วนนี้ นอกจากนี้เขียนชิ้นส่วนของโมเลกุล DNA ที่สิ่งนี้และ RNA ถูกสังเคราะห์ (สำหรับการใช้งานกับตารางของรหัสพันธุกรรม)
ตัวอย่างของภารกิจประเภทที่ห้า
- แฟรกเมนต์ DNA มีลำดับต่อไปนี้ของ Tatgggtstat Nucleotide ตั้งค่าลำดับนิวคลีโอไทด์ T-RNA ซึ่งสังเคราะห์จากส่วนนี้และกรดอะมิโนที่ T-RNA นี้จะถูกถ่ายโอนหาก Triplet ที่สามสอดคล้องกับ antiquodone t-rna เพื่อแก้ปัญหาให้ใช้ตารางรหัสพันธุกรรม
- แฟรกเมนต์ DNA มีลำดับต่อไปนี้ของ Nucleotide Cahagatttttt ต่อไปนี้ ตั้งค่าลำดับนิวคลีโอไทด์ T-RNA ซึ่งสังเคราะห์จากส่วนนี้และกรดอะมิโนที่ T-RNA นี้จะถูกถ่ายโอนหาก Triplet ที่สามสอดคล้องกับ antiquodone t-rna เพื่อแก้ปัญหาให้ใช้ตารางรหัสพันธุกรรม
- ชิ้นส่วน DNA มีลำดับต่อไปนี้ของ Nucleotide Hzzhattstg ตั้งค่าลำดับนิวคลีโอไทด์ T-RNA ซึ่งสังเคราะห์จากส่วนนี้และกรดอะมิโนที่ T-RNA นี้จะถูกถ่ายโอนหาก Triplet ที่สามสอดคล้องกับ antiquodone t-rna เพื่อแก้ปัญหาให้ใช้ตารางรหัสพันธุกรรม
- ส่วน DNA มีลำดับต่อไปนี้ของนิวคลีโอไทด์ tgtzhattsian ต่อไปนี้ ตั้งค่าลำดับนิวคลีโอไทด์ T-RNA ซึ่งสังเคราะห์จากส่วนนี้และกรดอะมิโนที่ T-RNA นี้จะถูกถ่ายโอนหาก Triplet ที่สามสอดคล้องกับ antiquodone t-rna เพื่อแก้ปัญหาให้ใช้ตารางรหัสพันธุกรรม
- ชิ้นส่วน DNA มีลำดับของ Zatgaategat Nucleotides ต่อไปนี้ ตั้งค่าลำดับนิวคลีโอไทด์ T-RNA ซึ่งสังเคราะห์จากส่วนนี้และกรดอะมิโนที่ T-RNA นี้จะถูกถ่ายโอนหาก Triplet ที่สามสอดคล้องกับ antiquodone t-rna เพื่อแก้ปัญหาให้ใช้ตารางรหัสพันธุกรรม
ตัวอย่างของภารกิจของประเภทที่หก
ตัวอย่างของงานประเภทที่เจ็ด
ภาคผนวก I รหัสพันธุกรรม (RNA)
| ฐานแรก | ฐานที่สอง | รากฐานที่สาม | |||
| ว. | ค. | แต่ | กรัม | ||
| ว. | เฟิน | เสร. | tir | cis | ว. |
| เฟิน | เสร. | tir | cis | ค. | |
| leu | เสร. | - | - | แต่ | |
| leu | เสร. | - | สาม | กรัม | |
| ค. | leu | มือโปร | gis | ขอร้อง | ว. |
| leu | มือโปร | gis | ขอร้อง | ค. | |
| leu | มือโปร | แวบ | ขอร้อง | แต่ | |
| leu | มือโปร | แวบ | ขอร้อง | กรัม | |
| แต่ | ไอล์ | tre | เอ็น | เสร. | ว. |
| ไอล์ | tre | เอ็น | เสร. | ค. | |
| ไอล์ | tre | การฉุนเฉียว | ขอร้อง | แต่ | |
| พบ | tre | การฉุนเฉียว | ขอร้อง | กรัม | |
| กรัม | เพลา | ala | งูเห่า | gly. | ว. |
| เพลา | ala | งูเห่า | gly. | ค. | |
| เพลา | ala | แดง | gly. | แต่ | |
| เพลา | ala | แดง | gly. | กรัม |
คำตอบ
- a \u003d. r \u003d c \u003d.
- a \u003d. r \u003d c \u003d.
- c \u003d. a \u003d t \u003d.
- c \u003d. a \u003d t \u003d.
- r \u003d. a \u003d t \u003d.
- r \u003d. a \u003d t \u003d.
- กรดอะมิโน, แฝดสามนิวคลีโอไทด์
- กรดอะมิโน, แฝดสามนิวคลีโอไทด์
- Triplet, กรดอะมิโน, โมเลกุล T-RNA
- Triplet กรดอะมิโนโมเลกุล T-RNA
- Triplets กรดอะมิโนโมเลกุล T-RNA
- และ RNA: Woods-GCC-CGA-HUC ลำดับกรดอะมิโน: Feng Ala-arg-Shaft
- และ RNA: GSU-AU-GHZ-TSUA ลำดับกรดอะมิโน: Gly-Ile-Glyhelu
- และ RNA: UCA-AAG-TSG-GUU ลำดับกรดอะมิโน: Ser-Liz-Pro-Shaft
- และ -RNA: Tsua-Aug-gau-caha ลำดับกรดอะมิโน: lei-met-asp-gln
- และ RNA: Gau-agg-cga-cag ลำดับกรดอะมิโน: ASP-arg-arg-gln
- และ rna: uuc-gau-huts ugh ลำดับกรดอะมิโน: FEN-ASP-Shaft-Three
- และ RNA: CCA-TSGG-CTSU-UUC ลำดับกรดอะมิโน: Pro-arg-Pro-Hondryer
- และ RNA: GGH-Cau-Uua-Agts ลำดับกรดอะมิโน: Gi-Gis-Lei-Ser
- ชิ้นส่วน DNA: tstaztzatgattt AntiCodones T-RNA: CSU, CSUC, Aug, AAG, ACHуу ลำดับกรดอะมิโน: ASP-GLU-TIR-FENG LIZ
- ชิ้นส่วน DNA: gcttztsayagggats AntiCodones T-RNA: GCU, CCA, UAA, GGG, ATSC ลำดับกรดอะมิโน: arg-gly-ile-pro-three
- ส่วน DNA: atsaagttattstststs AntiCodones T-RNA: ATS, AGU, UAU, CTSU, UZZ ลำดับกรดอะมิโน: CIS-Ser-Ile-Gly-arg
- ชิ้นส่วน DNA: ghtshttgtgtshtstch AntiCodones T-RNA: GHz, Guu, Gug, CGC, uuts ลำดับกรดอะมิโน: Pro-Gln Gis-Ala-Ser
- ชิ้นส่วน DNA: tgtzatsggtggg Anticodones T-RNA: Yuga, Tsgs, TSGG, UUG, GGA ลำดับกรดอะมิโน: Tre-Shaft-Ala-Asn-Pro
- ชิ้นส่วน DNA: tstgtctgagttsaga AntiCodones T-RNA: TSUG, Utsu, Gag, UUC, YEAH ลำดับกรดอะมิโน: ASP-arg lei-liz-ser
- ชิ้นส่วน DNA: azgtghattgtscat AntiCodones T-RNA: ACG, UGA, TSU, HCH, CAU ลำดับกรดอะมิโน: CIS-Tre-Depth Agr-Shaft
- ชิ้นส่วน DNA: Tsgtztsgstcatata AntiCodones T-RNA: CSU, CSG, Huts, AAU, AU ลำดับกรดอะมิโน: Ala-Gly-Gln Lei-tir
- ชิ้นส่วน DNA: Tsgattsaagahatg Anticodones T-RNA: CGA, Uua, CAA, GAA, ส.ค. ลำดับกรดอะมิโน: Ala-Asn Val-Lei-tir
- t-RNA: AUU CTSC-GAU-AAC anticodone gau, codon และ rna - csu, แบกกรดอะมิโน - lei
- t-RNA: Guu-Tsua-AAA-CAA Anticodone AAA, รหัสและ RNA - UU, การถ่ายโอนกรดอะมิโน - ไดร์เป่าผม
- t-RNA: TSGG-UU-AGG-ACU Antikodon AGG, Codon และ RNA - UZZZ, โอนกรดอะมิโน - Ser
- t-RNA: ACA-GSU-AGU -UUG Antikodon AGU รหัสและ RNA - UCA ถ่ายโอนกรดอะมิโน - Ser
- t-RNA: Gua-Tsuu-Uua-Tsua Anticodone Uua, รหัสและ RNA - AAU, กรดอะมิโน - ASN
- . ชุดพันธุกรรม:
- . ชุดพันธุกรรม:
- . ชุดพันธุกรรม:
- . ชุดพันธุกรรม:
- . ชุดพันธุกรรม:
- . ชุดพันธุกรรม:
- . ชุดพันธุกรรม:
- . ชุดพันธุกรรม:
- เนื่องจากโมเลกุลพีวีซีและ ATP เกิดขึ้นจากโมเลกุลกลูโคสหนึ่งตัวดังนั้น ATP จึงถูกสังเคราะห์ หลังจากขั้นตอนการใช้พลังงานของการสลายตัวโมเลกุล ATP (เมื่อโมเลกุลกลูโคสถูกแยกออก) ดังนั้น ATP จึงถูกสังเคราะห์ ผลรวมของการละลายคือ ATP
- เนื่องจากโมเลกุลพีวีซีและ ATP เกิดขึ้นจากโมเลกุลกลูโคสหนึ่งตัวดังนั้น ATP จึงถูกสังเคราะห์ หลังจากขั้นตอนการใช้พลังงานของการสลายตัวโมเลกุล ATP (เมื่อโมเลกุลกลูโคสถูกแยกออก) ดังนั้น ATP จึงถูกสังเคราะห์ ผลรวมของการละลายคือ ATP
- เนื่องจากโมเลกุลพีวีซีและ ATP เกิดขึ้นจากโมเลกุลกลูโคสหนึ่งตัวดังนั้น ATP จึงถูกสังเคราะห์ หลังจากขั้นตอนการใช้พลังงานของการสลายตัวโมเลกุล ATP (เมื่อโมเลกุลกลูโคสถูกแยกออก) ดังนั้น ATP จึงถูกสังเคราะห์ ผลรวมของการละลายคือ ATP
- โมเลกุลของ PVC เข้าสู่วงจร Krebs ดังนั้นโมเลกุลกลูโคสจึงเลิกกัน ปริมาณของ ATP หลังจาก glycolysis - โมเลกุลหลังจากช่วงพลังงาน - โมเลกุลผลรวมของการละลายของโมเลกุล ATP
- โมเลกุลของ PVC เข้าสู่วงจร Krebs ดังนั้นโมเลกุลกลูโคสจึงเลิกกัน ปริมาณของ ATP หลังจาก glycolysis - โมเลกุลหลังจากช่วงพลังงาน - โมเลกุลผลรวมของการละลายของโมเลกุล ATP
- โมเลกุลของ PVC เข้าสู่วงจร Krebs ดังนั้นโมเลกุลกลูโคสจึงเลิกกัน ปริมาณของ ATP หลังจาก glycolysis - โมเลกุลหลังจากช่วงพลังงาน - โมเลกุลผลรวมของการละลายของโมเลกุล ATP
- โมเลกุลของ PVC เข้าสู่วงจร Krebs ดังนั้นโมเลกุลกลูโคสจึงเลิกกัน ปริมาณของ ATP หลังจาก glycolysis - โมเลกุลหลังจากช่วงพลังงาน - โมเลกุลผลรวมของการละลายของโมเลกุล ATP
ทฤษฎีเซลล์บทบัญญัติหลักบทบาทในการก่อตัวของภาพวิทยาศาสตร์ธรรมชาติสมัยใหม่ของโลก การพัฒนาความรู้เกี่ยวกับเซลล์ โครงสร้างเซลลูล่าร์ของสิ่งมีชีวิตความคล้ายคลึงกันของโครงสร้างของเซลล์ของสิ่งมีชีวิตทั้งหมดเป็นพื้นฐานของความสามัคคีของโลกอินทรีย์หลักฐานของเครือข่ายสัตว์ป่า
กรง - หน่วยของโครงสร้างกิจกรรมที่สำคัญการเจริญเติบโตและการพัฒนาสิ่งมีชีวิต เซลล์ manifold ลักษณะเปรียบเทียบ เซลล์ของพืชสัตว์แบคทีเรียเห็ด
โครงสร้างของเซลล์ Pro- และ Eukaryotic ความสัมพันธ์ของโครงสร้างและหน้าที่ของชิ้นส่วนและเซลล์อิงเกราะเป็นพื้นฐานของความซื่อสัตย์ การเผาผลาญ: การแลกเปลี่ยนพลังงานและพลาสติกความสัมพันธ์ของพวกเขา เอนไซม์ลักษณะทางเคมีของพวกเขาบทบาทในการเผาผลาญ ขั้นตอนการแลกเปลี่ยนพลังงาน การหมักและการหายใจ การสังเคราะห์ด้วยแสงความหมายของมันบทบาทจักรวาล ขั้นตอนการสังเคราะห์แสง ปฏิกิริยาการสังเคราะห์แสงที่เบาและมืดความสัมพันธ์ของพวกเขา Chemosynthesis
I Protein Biosynthesis I. กรดนิวคลีอิก. ลักษณะของเมทริกซ์ธรรมชาติของปฏิกิริยาการสังเคราะห์การสังเคราะห์ ยีนรหัสพันธุกรรมและคุณสมบัติของมัน โครโมโซมโครงสร้างของพวกเขา (รูปร่างและขนาด) และฟังก์ชั่น จำนวนโครโมโซมและคงที่สปีชีส์ของพวกเขา การกำหนดชุดของโครโมโซมในเซลล์ร่างกายและอวัยวะเพศ วงจรชีวิตของเซลล์: interfac และ mitosis Mitoz - ส่วนของเซลล์ร่างกาย meiosis ขั้นตอน mitosis และ meiosis การพัฒนาเซลล์อวัยวะเพศในพืชและสัตว์ ความคล้ายคลึงกันและความแตกต่างระหว่าง mitosis และ meiosis ความหมายของพวกเขา การแบ่งเซลล์ - พื้นฐานของการเติบโตการพัฒนาและการสืบพันธุ์ของสิ่งมีชีวิต
กรงเป็นระบบชีวภาพ
ทฤษฎีมือถือสมัยใหม่บทบัญญัติหลักบทบาทในการก่อตัวของภาพวิทยาศาสตร์ธรรมชาติสมัยใหม่ของโลก การพัฒนาความรู้เกี่ยวกับเซลล์ โครงสร้างเซลลูล่าร์ของสิ่งมีชีวิต - พื้นฐานของความสามัคคีของโลกอินทรีย์หลักฐานของเครือข่ายสัตว์ป่า
ทฤษฎีเซลล์ที่ทันสมัยบทบัญญัติหลักของมันบทบาทในการก่อตัวของภาพวิทยาศาสตร์ธรรมชาติที่ทันสมัยของโลก
หนึ่งในแนวคิดพื้นฐานในชีววิทยาสมัยใหม่คือความคิดที่ว่าโครงสร้างมือถือมีอยู่ในทุกสิ่งมีชีวิต การศึกษาโครงสร้างของเซลล์การดำรงชีวิตและการมีปฏิสัมพันธ์กับสิ่งแวดล้อม เซลล์วิทยาขณะนี้มักจะเรียกว่าชีววิทยาของเซลล์มากขึ้น เซลล์วิทยามีหน้าที่กำหนดทฤษฎีเซลล์ (1838-1839, M. Shleden, T. Svann เสริมในปี 1855 R. Virhov)
ทฤษฎีเชลเลน มันเป็นแนวคิดทั่วไปของโครงสร้างและหน้าที่ของเซลล์เป็นมหาวิทยาลัยเกี่ยวกับการสืบพันธุ์และบทบาทของพวกเขาในการก่อตัวของสิ่งมีชีวิตหลายเซลล์
บทบัญญัติหลักของทฤษฎีเซลล์:
- เซลล์ - หน่วยของโครงสร้างกิจกรรมที่สำคัญการเจริญเติบโตและการพัฒนาสิ่งมีชีวิต - ไม่มีชีวิตของชีวิต
- เซลล์เป็นระบบเดียวที่ประกอบด้วยองค์ประกอบที่หลากหลายเชื่อมต่อกันตามธรรมชาติซึ่งเป็นการศึกษาแบบองค์รวม
- เซลล์ของสิ่งมีชีวิตทั้งหมดมีความคล้ายคลึงกับองค์ประกอบทางเคมีโครงสร้างและฟังก์ชั่น
- เซลล์ใหม่จะเกิดขึ้นจากการแบ่งเซลล์ของมารดา ("เซลล์จากเซลล์")
- เซลล์ของสิ่งมีชีวิตหลายเซลล์รูปแบบของเนื้อเยื่ออวัยวะประกอบด้วยเนื้อเยื่อ ชีวิตของร่างกายโดยรวมเป็นเพราะการมีปฏิสัมพันธ์ของส่วนประกอบของเซลล์
- เซลล์ของสิ่งมีชีวิตหลายเซลล์มียีนที่สมบูรณ์ แต่แตกต่างกันไปในแต่ละกลุ่มที่พวกเขามีกลุ่มที่แตกต่างกันของยีนผลที่ตามมาซึ่งเป็นความหลากหลายทางสัณฐานวิทยาและการทำงานของเซลล์ - ความแตกต่าง
ด้วยการสร้างทฤษฎีเซลล์มันชัดเจนว่าเซลล์เป็นหน่วยที่เล็กที่สุดของชีวิตระบบการใช้ชีวิตระดับประถมศึกษาซึ่งมีอยู่ในสัญญาณทั้งหมดและคุณสมบัติของการใช้ชีวิตทั้งหมด การกำหนดทฤษฎีเซลล์ได้กลายเป็นข้อกำหนดเบื้องต้นที่สำคัญที่สุดสำหรับการพัฒนามุมมองเกี่ยวกับการถ่ายทอดทางพันธุกรรมและความแปรปรวนเนื่องจากการระบุลักษณะของพวกเขาและรูปแบบที่มีอยู่ในนั้นอย่างหลีกเลี่ยงไม่ได้แนะนำแนวคิดของความเป็นสากลของโครงสร้างของสิ่งมีชีวิต การระบุความสามัคคีขององค์ประกอบทางเคมีและแผนโครงสร้างของเซลล์คือแรงผลักดันสำหรับการพัฒนาความคิดเกี่ยวกับที่มาของสิ่งมีชีวิตและวิวัฒนาการของพวกเขา นอกจากนี้ที่มาของสิ่งมีชีวิตหลายเซลล์จากเซลล์เดียวในกระบวนการของการพัฒนาตัวอ่อนเป็นความเชื่อของตัวอ่อนที่ทันสมัย
การพัฒนาความรู้เกี่ยวกับเซลล์
จนกระทั่งศตวรรษที่ XVII คนไม่ทราบอะไรเลยเกี่ยวกับโครงสร้างจุลภาคของรายการโดยรอบและรับรู้โลกด้วยตาเปล่า อุปกรณ์สำหรับการศึกษา Microman - กล้องจุลทรรศน์ถูกคิดค้นโดยประมาณในปี 1590 โดยกลศาสตร์ชาวดัตช์ G. และ Z. Jansen อย่างไรก็ตามความไม่สมบูรณ์ของเขาไม่ได้ให้โอกาสในการพิจารณาวัตถุขนาดเล็กเพียงพอ เฉพาะการสร้างบนพื้นฐานของกล้องจุลทรรศน์ที่ซับซ้อนที่เรียกว่า K. Durbel (1572-1634) สนับสนุนความคืบหน้าในพื้นที่นี้
ในปี 1665 นักวิทยาศาสตร์นักวิทยาศาสตร์ชาวอังกฤษ R. Guk (1635-1703) ปรับปรุงการออกแบบของกล้องจุลทรรศน์และเทคโนโลยีการบดเลนส์และต้องการให้แน่ใจว่าในการปรับปรุงคุณภาพของภาพตรวจสอบการตัดถ่านและพืชที่อยู่อาศัยภายใต้มัน บนบาดแผลเขาค้นพบรูขุมขนที่เล็กที่สุดคล้ายรังผึ้งผึ้งและเรียกพวกเขาว่าเซลล์ (จาก lat. เซลลา - เซลล์กรง) เป็นที่น่าสนใจที่จะทราบว่า R. GUK พิจารณาองค์ประกอบหลักของ Cellular Cage
ในช่วงครึ่งหลังของศตวรรษที่ XVII งานของกล้องจุลทรรศน์ที่โดดเด่นที่สุด M. Malpigi (1628-1694) และ N. เพิ่มขึ้น (1641-1712) นอกจากนี้ยังพบโครงสร้างมือถือของพืชจำนวนมาก
เพื่อให้แน่ใจว่าเขาเห็น R. หนาและนักวิทยาศาสตร์คนอื่น ๆ คือความจริงที่ไม่มีการก่อตัวเป็นพิเศษของพ่อค้าชาวดัตช์ A. Van Levenguk ได้พัฒนาการออกแบบกล้องจุลทรรศน์ที่แตกต่างจากที่มีอยู่แล้วและปรับปรุงเทคโนโลยี ของเลนส์การผลิต สิ่งนี้ทำให้เขาสามารถเพิ่มขึ้น 275-300 ครั้งและพิจารณารายละเอียดดังกล่าวของโครงสร้างที่ไม่สามารถใช้งานได้ทางเทคนิคจากนักวิทยาศาสตร์ที่เหลือ A. Van Levenguk เป็นผู้สังเกตการณ์ที่ไม่มีใครเทียบ: เขาสเก็ตช์อย่างรอบคอบและอธิบายสิ่งที่เห็นภายใต้กล้องจุลทรรศน์ แต่ไม่ได้พยายามอธิบาย มันเปิดสิ่งมีชีวิตเซลล์เดียวรวมถึงแบคทีเรียในเซลล์ของพืชเมล็ดพบคลอโรพลาสต์หนาของผนังเซลล์ แต่เป็นไปได้ที่จะประเมินการค้นพบของมันมากขึ้นในภายหลัง
การเปิดตัวขององค์ประกอบของโครงสร้างภายในของสิ่งมีชีวิตในช่วงครึ่งแรกของศตวรรษที่ XIX ติดตามหนึ่งหลังจากนั้น G. mole โดดเด่นในเซลล์พืชสารสดและของเหลวน้ำ - น้ำผลไม้เซลลูล่าร์ค้นพบรูขุมขน นักพฤกษศาสตร์อังกฤษ R. Brown (1773-1858) ในปี 1831 เปิดเคอร์เนลในเซลล์กล้วยไม้แล้วพบได้ในทุกเซลล์พืช Czech Scholar Ya Purkinje (1787-1869) เพื่อแสดงถึงศูนย์กึ่งของเหลวของเซลล์คำว่า "Protoplasm" (1840) ได้รับการแนะนำ ด้านหลังโคตรนักพฤกษศาสตร์ชาวเบลเยียมเอ็ม Shleden (1804-1881) ขั้นสูงซึ่งศึกษาการพัฒนาและความแตกต่างของโครงสร้างเซลลูลาร์ที่หลากหลายของพืชที่สูงขึ้นพิสูจน์ให้เห็นว่าสิ่งมีชีวิตพืชทั้งหมดนำไปสู่ต้นกำเนิดจากเซลล์หนึ่ง นอกจากนี้เขายังพิจารณาในเมล็ดของเซลล์สาธารณรัฐเช็กไหล่โค้งมนของนิวเคลียส (1842)
ในปี 1827 ตัวกมกรัสเซีย K. Baer ค้นพบไข่ของไข่และสัตว์เลี้ยงลูกด้วยนมอื่น ๆ จึงมีความคิดเกี่ยวกับการพัฒนาของร่างกายโดยเฉพาะจากน้ำหนักของผู้ชาย นอกจากนี้เขายังพิสูจน์การก่อตัวของสัตว์หลายเซลล์ของร่างกายจากเซลล์เดียว - ไข่ที่ปฏิสนธิเช่นเดียวกับความคล้ายคลึงกันของขั้นตอนของการพัฒนาเชื้อโรคของสัตว์หลายเซลล์ซึ่งได้พิสูจน์แล้วว่าเป็นความคิดของความสามัคคี ของที่มาของพวกเขา ข้อมูลที่สะสมอยู่ตรงกลางของศตวรรษที่ XIX เรียกร้องโดยทั่วไปซึ่งทฤษฎีเซลล์กลายเป็น ชีววิทยาสูตรของมันมีภาระผูกพันกับนักสัตววิทยาชาวเยอรมัน T. Schvanna (1810-1882) ซึ่งบนพื้นฐานของข้อมูลและข้อสรุปของตัวเอง M. Shleiden ในการพัฒนาของพืชหยิบยกข้อสันนิษฐานว่าถ้าแกนอยู่ในใด ๆ การก่อตัวภายใต้กล้องจุลทรรศน์การศึกษานี้เป็นกรง ขึ้นอยู่กับเกณฑ์นี้ T. Svann กำหนดตำแหน่งหลักของทฤษฎีเซลล์
แพทย์ชาวเยอรมันและนักพยาธิวิทยา R. Virhov (1821-1902) ได้เปิดตัวตำแหน่งที่สำคัญอีกประการหนึ่งในทฤษฎีนี้: เซลล์เกิดขึ้นโดยการหารเซลล์เริ่มต้น I.e. เซลล์จะเกิดขึ้นจากเซลล์ ("เซลล์จากเซลล์" เท่านั้น
ตั้งแต่การสร้างทฤษฎีเซลล์หลักคำสอนของเซลล์เป็นหน่วยของโครงสร้างฟังก์ชั่นและการพัฒนาของร่างกายได้รับการพัฒนาอย่างต่อเนื่อง ในตอนท้ายของศตวรรษที่สิบเก้าเนื่องจากความสำเร็จของเทคโนโลยีกล้องจุลทรรศน์โครงสร้างของเซลล์ได้รับการระบุว่ามีการอธิบายอวัยวะ - ชิ้นส่วนของเซลล์ที่มีฟังก์ชั่นต่าง ๆ วิธีการก่อตัวของเซลล์ใหม่ (mitosis, meysosis) และ กลายเป็นมูลค่าหลักของโครงสร้างเซลล์ในการถ่ายโอนคุณสมบัติทางพันธุกรรม การใช้วิธีการวิจัยทางเคมีกายภาพล่าสุดทำให้เป็นไปได้ที่จะลึกลงไปในการจัดเก็บข้อมูลและการส่งข้อมูลทางพันธุกรรมรวมถึงการสำรวจโครงสร้างที่ละเอียดอ่อนของแต่ละโครงสร้างเซลล์ ทั้งหมดนี้มีส่วนทำให้ไฮไลต์ของวิทยาศาสตร์ของเซลล์เป็นสาขาอิสระของความรู้ เซลล์วิทยา.
โครงสร้างเซลลูล่าร์ของสิ่งมีชีวิตความคล้ายคลึงกันของโครงสร้างของเซลล์ของสิ่งมีชีวิตทั้งหมดเป็นพื้นฐานของความสามัคคีของโลกอินทรีย์หลักฐานของเครือข่ายสัตว์ป่า
สิ่งมีชีวิตทั้งหมดที่รู้จักกันในวันที่ (พืชสัตว์เห็ดและแบคทีเรีย) มีโครงสร้างมือถือ แม้แต่ไวรัสที่ไม่มีโครงสร้างมือถือสามารถคูณได้เฉพาะในเซลล์ เซลล์เป็นหน่วยงานที่ใช้งานอยู่ในโครงสร้างพื้นฐานของการใช้ชีวิตซึ่งมีอยู่ในลักษณะทั้งหมดโดยเฉพาะอย่างยิ่งการเผาผลาญและการเปลี่ยนแปลงของพลังงาน Homeostasis การเจริญเติบโตและการพัฒนาการสืบพันธุ์และหงุดหงิด ในขณะเดียวกันก็เก็บไว้ในเซลล์ข้อมูลทางพันธุกรรมจะถูกประมวลผลและดำเนินการ
แม้จะมีทุกเซลล์ที่หลากหลายโครงสร้างของโครงสร้างเป็นหนึ่งเดียว: พวกเขาทั้งหมดมี อุปกรณ์ที่มีสุขภาพดีแช่โดย B. พลาสซึมและเซลล์โดยรอบ เมมเบรนพลาสม่า.
เซลล์เกิดขึ้นจากวิวัฒนาการในระยะยาวของโลกอินทรีย์ การรวมกันของเซลล์ในสิ่งมีชีวิตหลายเซลล์ไม่ใช่การรวมอย่างง่ายเนื่องจากแต่ละเซลล์ในขณะที่ยังคงรักษาสัญญาณทั้งหมดที่มีอยู่ในสิ่งมีชีวิตในเวลาเดียวกันได้รับคุณสมบัติใหม่เนื่องจากการปฏิบัติตามฟังก์ชั่นเฉพาะ ในมือข้างหนึ่งร่างกายหลายเซลล์สามารถแบ่งออกเป็นส่วนประกอบของชิ้นส่วนของชิ้นส่วน - เซลล์ แต่ในทางกลับกันพับได้อีกครั้งมันเป็นไปไม่ได้ที่จะเรียกคืนฟังก์ชั่นของสิ่งมีชีวิตแบบองค์รวมเนื่องจากคุณสมบัติใหม่ปรากฏในปฏิสัมพันธ์ของ บางส่วนของระบบ นี่เป็นที่ประจักษ์โดยหนึ่งในรูปแบบพื้นฐานที่มีลักษณะการใช้ชีวิตความสามัคคีของการแยกและองค์รวม ขนาดเล็กและเซลล์จำนวนมากสร้างพื้นผิวขนาดใหญ่ที่จำเป็นในสิ่งมีชีวิตหลายเซลล์ที่จำเป็นเพื่อให้แน่ใจว่าการเผาผลาญอย่างรวดเร็ว นอกจากนี้ในกรณีของการตายของส่วนหนึ่งของร่างกายความสมบูรณ์ของมันสามารถคืนค่าได้โดยการสืบพันธุ์ของเซลล์ นอกเซลล์ที่เก็บข้อมูลและการถ่ายโอนข้อมูลทางพันธุกรรมการจัดเก็บและการถ่ายโอนพลังงานตามด้วยการเปิดใช้งาน ในที่สุดการแยกการทำงานระหว่างเซลล์ในร่างกายหลายเซลล์ทำให้มั่นใจได้ถึงความเป็นไปได้ที่กว้างขวางของการปรับตัวเข้ากับที่อยู่อาศัยและปรากฏคำบุพบทขององค์กรของพวกเขา
ดังนั้นการจัดตั้งความสามัคคีของแผนสำหรับโครงสร้างของเซลล์ของสิ่งมีชีวิตทั้งหมดคือการพิสูจน์ความสามัคคีของต้นกำเนิดของการใช้ชีวิตทั้งหมดบนโลก
เซลล์ manifold เซลล์ ProCarniotic และ Eukaryotic ลักษณะเปรียบเทียบของพืชสัตว์เชื้อแบคทีเรียเห็ดพันธุ์เซลล์
ตามทฤษฎีเซลล์ของเซลล์เป็นหน่วยโครงสร้างที่เล็กที่สุดและการทำงานของสิ่งมีชีวิตซึ่งมีอยู่ในคุณสมบัติทั้งหมดของการใช้ชีวิต ด้วยจำนวนของเซลล์สิ่งมีชีวิตแบ่งออกเป็นเซลล์เดียวและหลายเซลล์ เซลล์ของสิ่งมีชีวิตเซลล์เดียวมีอยู่เป็นสิ่งมีชีวิตอิสระและดำเนินการทุกฟังก์ชั่นของการใช้ชีวิต Unicellularity เป็น Prokaryotes ทั้งหมดและ Eukaryotes จำนวนหนึ่ง (Algae หลายประเภท, เห็ดและสัตว์ที่ง่ายที่สุด) ซึ่งโดดเด่นในรูปทรงและขนาดที่หลากหลาย อย่างไรก็ตามสิ่งมีชีวิตส่วนใหญ่ยังคงเป็นหลายเซลล์ เซลล์ของพวกเขามีความเชี่ยวชาญในการทำหน้าที่บางอย่างและรูปแบบเนื้อเยื่อและอวัยวะซึ่งไม่สามารถสะท้อนถึงคุณสมบัติทางสัณฐานวิทยา ตัวอย่างเช่นร่างกายมนุษย์เกิดขึ้นจากประมาณ 10 14 เซลล์ที่แสดงโดยประมาณ 200 สายพันธุ์ที่มีหลากหลายรูปแบบและขนาด
รูปร่างของเซลล์สามารถโค้งมน, ทรงกระบอก, ลูกบาศก์, ปริซึม, diskidal, beltidoid, ดาว, ฯลฯ และดาว - เซลล์ของเนื้อเยื่อประสาท จำนวนของเซลล์ไม่มีรูปแบบคงที่เลย เหล่านี้รวมถึงก่อนอื่นเม็ดเลือดแดงเลือด
ขนาดของเซลล์ยังแตกต่างกันอย่างมีนัยสำคัญ: เซลล์ส่วนใหญ่ของสิ่งมีชีวิตหลายเซลล์มีขนาดตั้งแต่ 10 ถึง 100 μmและเล็กที่สุด - 2-4 ไมครอน ขีด จำกัด ที่ต่ำกว่าเกิดจากความจริงที่ว่าเซลล์ต้องมีชุดของสารและโครงสร้างขั้นต่ำเพื่อให้แน่ใจว่ากิจกรรมที่สำคัญและขนาดของเซลล์ขนาดใหญ่เกินไปจะป้องกันการแลกเปลี่ยนสารและพลังงานกับสภาพแวดล้อมและจะทำให้มันยากสำหรับการบำรุงรักษา homeostasis อย่างไรก็ตามบางเซลล์สามารถดูได้ด้วยตาเปล่า ก่อนอื่นมันรวมถึงเซลล์ของผลไม้ของแตงโมและต้นแอปเปิ้ลเช่นเดียวกับไข่ปลาและนก แม้ว่าหนึ่งในขนาดของเซลล์เชิงเส้นเกินตัวบ่งชี้เฉลี่ยอื่น ๆ ทั้งหมดที่สอดคล้องกับบรรทัดฐาน ตัวอย่างเช่นการดำเนินคดีเซลล์ประสาทสามารถยาวเกิน 1 เมตร แต่เส้นผ่านศูนย์กลางของมันจะยังคงสอดคล้องกับค่าเฉลี่ย ไม่มีการพึ่งพาโดยตรงระหว่างขนาดเซลล์และขนาดร่างกาย ดังนั้นเซลล์ของกล้ามเนื้อของช้างและหนูมีมิติเท่ากัน
เซลล์ Prokaryotic และ Eukaryotic
ดังที่กล่าวไว้ข้างต้นเซลล์มีคุณสมบัติการทำงานที่คล้ายคลึงกันมากมายและคุณสมบัติทางสัณฐานวิทยา แต่ละห้องประกอบด้วยไซโตพลาสซึมที่แช่อยู่ในนั้น เครื่องมือทางพันธุกรรมและแยกออกจากสภาพแวดล้อมภายนอก เมมเบรนพลาสม่า, หรือ พลาสมาล์มไม่ขัดขวางการประมวลผลการเผาผลาญและพลังงาน ด้านนอกของเมมเบรนที่เซลล์สามารถเป็นผนังเซลล์อื่นที่ประกอบด้วยสารต่าง ๆ ซึ่งทำหน้าที่ปกป้องเซลล์และเป็นโครงกระดูกภายนอก
ไซโตพลาสซึมเป็นเนื้อหาทั้งหมดของเซลล์ที่เติมช่องว่างระหว่างพลาสมาเมมเบรนและโครงสร้างที่มีข้อมูลทางพันธุกรรม ประกอบด้วยสารหลัก - galoplasma - และช่องเติมเงินและการรวมเข้ากันอยู่ในนั้น organoid - เหล่านี้เป็นส่วนประกอบถาวรของเซลล์ที่ทำหน้าที่บางอย่างและการรวม - เซลล์ที่หายไปในช่วงชีวิตของส่วนประกอบของเซลล์ที่ทำหน้าที่อะไหล่หรือขับถ่ายเป็นหลัก บ่อยครั้งที่การรวมแบ่งออกเป็นของแข็งและของเหลว การรวมที่เป็นของแข็งจะแสดงเป็นหลักโดยเม็ดและอาจมีลักษณะที่แตกต่างกันในขณะที่ vacuoles และหยดไขมันถือเป็นสิ่งที่มีการรวมของเหลว

ปัจจุบันแยกความแตกต่างระหว่างสองประเภทหลักขององค์กรหลัก: Prokaryotic และ Eukaryotic
เซลล์ prokaryotic ไม่มีนิวเคลียสข้อมูลทางพันธุกรรมไม่ได้แยกออกจากไซโตพลาสซึมกับเยื่อหุ้มเซลล์
พื้นที่ของไซโตพลาสซึมที่ข้อมูลทางพันธุกรรมถูกเก็บไว้ในเซลล์ prokaryotic ที่เรียกว่า นิวเคลียส. ในไซโตพลาสซึมของเซลล์ prokaryotic มีส่วนใหญ่เป็นหนึ่งประเภทของ organoid - Ribosomes และไม่มีเยื่อหุ้มอินทรีย์เลย Prokaryotm เป็นแบคทีเรีย
เซลล์ยูคาริโอต - เซลล์ที่อย่างน้อยหนึ่งขั้นตอนของการพัฒนามี แกนกลาง - โครงสร้างพิเศษที่ดีเอ็นเอตั้งอยู่
ไซโตพลาสซึมของเซลล์ยูคาริโอตนั้นแตกต่างจากความหลากหลายของเมมเบรนและอริกบอร์ดที่ไม่ใช่สัญลักษณ์ สิ่งมีชีวิต Eukaryotic รวมถึงพืชสัตว์และเห็ด มิติของเซลล์ prokaryotic นั้นเป็นกฎคำสั่งของขนาดที่น้อยกว่าขนาดของยูคาริโอต Pricar ส่วนใหญ่เป็นสิ่งมีชีวิต Unicellular และ Eukaryotes เป็น MultiCellular
ลักษณะเปรียบเทียบของโครงสร้างของพืชสัตว์แบคทีเรียและเห็ด
นอกเหนือจากลักษณะของ prokaryotes และยูคาริโอท, พืช, เซลล์สัตว์, สัตว์, เห็ดและแบคทีเรียมีคุณสมบัติอีกมากมาย ดังนั้นเซลล์พืชจึงมีช่องว่างเฉพาะ - คลอโรพลาสต์ซึ่งกำหนดความสามารถในการสังเคราะห์ด้วยแสงในขณะที่สิ่งมีชีวิตอื่นไม่เป็นไปตามสิ่งมีชีวิตเหล่านี้ แน่นอนว่านี่ไม่ได้หมายความว่าสิ่งมีชีวิตอื่น ๆ ไม่สามารถสังเคราะห์ด้วยแสงได้เช่นโดยแบคทีเรียมันจะไหลบนพลาสม่าม่าและฟองเมมเบรนแต่ละตัวในไซโตพลาสซึม
เซลล์ผักตามกฎมีฉะคลำขนาดใหญ่ที่เต็มไปด้วยน้ำผลไม้เซลลูลาร์ ในเซลล์สัตว์เชื้อราและแบคทีเรียพวกเขาก็พบ แต่มีต้นกำเนิดที่แตกต่างอย่างสิ้นเชิงและทำหน้าที่อื่น ๆ สารอะไหล่หลักที่เกิดขึ้นในรูปแบบของการรวมที่เป็นของแข็งในพืชเป็นแป้งในสัตว์และเห็ด - ไกลโคเจนและในแบคทีเรีย - ไกลโคเจนหรือ Volutin
อีกคุณสมบัติที่โดดเด่นของกลุ่มสิ่งมีชีวิตเหล่านี้คือองค์กรของอุปกรณ์พื้นผิว: ในเซลล์ของสิ่งมีชีวิตสัตว์ไม่มีผนังเซลล์เมมเบรนพลาสม่าของพวกเขาถูกปกคลุมด้วย glycocalix บาง ๆ ในขณะที่คนอื่นมีอยู่ สิ่งนี้อธิบายทั้งหมดเนื่องจากวิธีการให้อาหารสัตว์มีความเกี่ยวข้องกับการจับภาพอนุภาคอาหารในกระบวนการ Phagocytosis และการปรากฏตัวของผนังเซลล์จะกีดกันพวกเขาถึงความเป็นไปได้นี้ ลักษณะทางเคมีของสารที่รวมอยู่ในผนังเซลล์ไม่เหมือนกันในกลุ่มสิ่งมีชีวิตในกลุ่มต่าง ๆ : หากมีพืชเซลลูโลสจากนั้นเชื้อราคือไคตินและแบคทีเรีย - Marein ลักษณะเปรียบเทียบของโครงสร้างของพืชสัตว์เห็ดและแบคทีเรีย
| เข้าสู่ระบบ | แบคทีเรีย | สัตว์ | เห็ด | พืช |
| วิธีการโภชนาการ | heterotrophic หรือ autotrophic | heterotrophic | heterotrophic | avtotrophny |
| องค์กรของข้อมูลทางพันธุกรรม | Procarniot | eukaryota | eukaryota | eukaryota |
| สถานที่ดีเอ็นเอ | นิวเคลียสพลาสมิด | หลัก, mitochondria | หลัก, mitochondria | core, mitochondria, plastids |
| เมมเบรนพลาสม่า | มี | มี | มี | มี |
| ผนังเซลล์ | mareinova | — | Chitinova | เกี่ยวกับเซลลูโลส |
| พลาสซึม | มี | มี | มี | มี |
| organoid | ไรโบโซม | เมมเบรนและไม่ลักลอบนำเข้ารวมถึงศูนย์บริการ | เมมเบรนและไม่ทุบ | เมมเบรนและไม่ลักลอบนำเข้ารวมถึงผู้ส่งเพลิง |
| อวัยวะในการเคลื่อนไหว | Flagella และ Valli | Flagella และ Cilia | Flagella และ Cilia | Flagella และ Cilia |
| vacuole | ไม่ค่อย | ตัดย่อยอาหาร | บางครั้ง | Vacuole กลางที่มีน้ำผลไม้เซลลูลาร์ |
| การรวม | ไกลโคเจน, volusutin | ไกลโคเจน | ไกลโคเจน | แป้ง |
ความแตกต่างในโครงสร้างของเซลล์ของตัวแทนของอาณาจักรที่แตกต่างกันของสัตว์ป่าจะแสดงในรูป

องค์ประกอบทางเคมีของเซลล์ องค์ประกอบมาโครและการติดตาม ความสัมพันธ์ระหว่างโครงสร้างและหน้าที่ของสารอนินทรีย์และสารอินทรีย์ (โปรตีน, กรดนิวคลีอิก, คาร์โบไฮเดรต, ไขมัน lipids) ซึ่งรวมอยู่ในเซลล์ บทบาทของสารเคมีในเซลล์และร่างกายมนุษย์
องค์ประกอบทางเคมีของเซลล์
ในฐานะที่เป็นส่วนหนึ่งของสิ่งมีชีวิตองค์ประกอบทางเคมีส่วนใหญ่ของระบบธาตุต่าง ๆ ของ Elements Di Mendeleev เปิดจนถึงวันที่ถูกค้นพบ ในมือข้างหนึ่งพวกเขาไม่มีองค์ประกอบเดียวที่จะไม่อยู่ในลักษณะที่ไม่มีชีวิต แต่ในทางกลับกันความเข้มข้นของพวกเขาในร่างกายของธรรมชาติที่ไม่มีชีวิตและสิ่งมีชีวิตแตกต่างกันอย่างมีนัยสำคัญ
องค์ประกอบทางเคมีเหล่านี้เป็นสารอนินทรีย์และสารอินทรีย์ แม้จะมีความจริงที่ว่าสารอนินทรีย์มีอิทธิพลเหนือสิ่งมีชีวิต แต่เป็นสารอินทรีย์ที่กำหนดเอกลักษณ์ขององค์ประกอบทางเคมีและปรากฏการณ์ของชีวิตโดยรวมเนื่องจากพวกเขาถูกสังเคราะห์โดยสิ่งมีชีวิตในกระบวนการของชีวิตและการเล่นในปฏิกิริยามากที่สุด บทบาทสำคัญ.

การศึกษาองค์ประกอบทางเคมีของสิ่งมีชีวิตและปฏิกิริยาเคมีที่เกิดขึ้นในพวกเขาโดยวิทยาศาสตร์ ชีวเคมี.
ควรสังเกตว่าเนื้อหาของสารเคมีในเซลล์และเนื้อเยื่อต่าง ๆ อาจแตกต่างกันไปได้อย่างมีนัยสำคัญ ตัวอย่างเช่นหากโปรตีนมีอิทธิพลเหนือสารประกอบอินทรีย์ในเซลล์สัตว์จากนั้นในเซลล์ของพืช - คาร์โบไฮเดรต
| องค์ประกอบทางเคมี | เปลือกโลก | น้ำทะเล | สิ่งมีชีวิต |
| O. | 49.2 | 85.8 | 65-75 |
| ค. | 0.4 | 0.0035 | 15-18 |
| เอช. | 1.0 | 10.67 | 8-10 |
| น. | 0.04 | 0.37 | 1.5-3.0 |
| พี. | 0.1 | 0.003 | 0.20-1.0 |
| S. | 0.15 | 0.09 | 0.15-0.2 |
| เค. | 2.35 | 0.04 | 0.15-0.4 |
| แคลิฟอร์เนีย | 3.25 | 0.05 | 0.04-2.0 |
| cl. | 0.2 | 0.06 | 0.05-0.1 |
| mg. | 2.35 | 0.14 | 0.02-0.03 |
| นา. | 2.4 | 1.14 | 0.02-0.03 |
| fe | 4.2 | 0.00015 | 0.01-0.015 |
| zn. | < 0.01 | 0.00015 | 0.0003 |
| cu. | < 0.01 | < 0.00001 | 0.0002 |
| ผม. | < 0.01 | 0.000015 | 0.0001 |
| F. | 0.1 | 2.07 | 0.0001 |
มาโครและไมโครเซลล์
ในสิ่งมีชีวิตมีประมาณ 80 องค์ประกอบทางเคมี แต่มีเพียง 27 ขององค์ประกอบเหล่านี้ฟังก์ชั่นของพวกเขาในเซลล์และร่างกายได้รับการติดตั้ง องค์ประกอบที่เหลืออยู่ในปริมาณเล็กน้อยและเห็นได้ชัดว่าตกอยู่ในร่างกายด้วยอาหารน้ำและอากาศ เนื้อหาขององค์ประกอบทางเคมีในร่างกายแตกต่างกันอย่างมีนัยสำคัญ พวกเขาแบ่งออกเป็นองค์ประกอบ macroelements และการติดตาม
ความเข้มข้นของแต่ละ macroelements ร่างกายเกิน 0.01% และเนื้อหาทั้งหมดของพวกเขาคือ 99% Macroelements ได้แก่ ออกซิเจน, คาร์บอน, ไฮโดรเจน, ไนโตรเจน, ฟอสฟอรัส, ซัลเฟอร์, โพแทสเซียม, แคลเซียม, โซเดียม, คลอรีน, แมกนีเซียมและเหล็ก สี่องค์ประกอบแรกขององค์ประกอบ (ออกซิเจน, คาร์บอน, ไฮโดรเจนและไนโตรเจน) เรียกอีกอย่างว่า เกี่ยวกับ Organogenicเนื่องจากเป็นส่วนหนึ่งของสารประกอบอินทรีย์หลัก ฟอสฟอรัสและซัลเฟอร์เป็นส่วนประกอบของสารอินทรีย์จำนวนหนึ่งเช่นโปรตีนและกรดนิวคลีอิก ฟอสฟอรัสเป็นสิ่งจำเป็นสำหรับการก่อตัวของกระดูกและฟัน
หากไม่มี meCroelements ที่เหลืออยู่การทำงานปกติของร่างกายเป็นไปไม่ได้ ดังนั้นโพแทสเซียมโซเดียมและคลอรีนมีส่วนร่วมในกระบวนการกระตุ้นเซลล์ โพแทสเซียมยังจำเป็นสำหรับการทำงานของเอนไซม์และการเก็บรักษาน้ำในเซลล์ แคลเซียมเป็นส่วนหนึ่งของผนังเซลล์ของพืชกระดูกฟันและเปลือกหอยและจำเป็นต้องลดเซลล์กล้ามเนื้อรวมถึงการเคลื่อนไหวของเซลล์ แมกนีเซียมเป็นส่วนประกอบของเม็ดสีคลอโรฟิลล์ที่ช่วยให้การไหลเวียนของการสังเคราะห์แสง นอกจากนี้เขายังมีส่วนร่วมในการสังเคราะห์โปรตีน เหล็กนอกเหนือไปจากความจริงที่ว่ามันเป็นส่วนหนึ่งของฮีโมโกลบินที่ถือออกซิเจนในเลือดมันเป็นสิ่งจำเป็นสำหรับการไหลของระบบทางเดินหายใจและการสังเคราะห์แสงเช่นเดียวกับการทำงานของเอนไซม์จำนวนมาก
ไมโครเซลล์ มันมีอยู่ในร่างกายในความเข้มข้นน้อยกว่า 0.01% และความเข้มข้นทั้งหมดของพวกเขาในเซลล์ไม่ถึง 0.1% Microelements รวมถึงสังกะสีทองแดงแมงกานีสโคบอลต์ไอโอดีนฟลูออรีน ฯลฯ สังกะสีเป็นส่วนหนึ่งของโมเลกุลฮอร์โมนตับอ่อน - อินซูลินทองแดงเป็นสิ่งจำเป็นสำหรับการสังเคราะห์ด้วยแสงและการหายใจ โคบอลต์เป็นส่วนประกอบของวิตามินบี 12 การขาดซึ่งนำไปสู่โรคโลหิตจาง ไอโอดีนเป็นสิ่งจำเป็นสำหรับการสังเคราะห์ฮอร์โมนต่อมไทรอยด์ให้การเผาผลาญปกติและฟลูออรีนมีความเกี่ยวข้องกับการก่อตัวของเคลือบฟันของฟัน
ทั้งข้อบกพร่องและส่วนเกินหรือการละเมิดของการแลกเปลี่ยนองค์ประกอบมาโครและการติดตามนำไปสู่การพัฒนาโรคต่าง ๆ โดยเฉพาะอย่างยิ่งการขาดแคลเซียมและฟอสฟอรัสทำให้เกิดโรคกระดูกอ่อนการขาดไนโตรเจน - โปรตีนที่หนักหนาล้มเหลวการขาดธาตุเหล็ก - โรคโลหิตจางและการขาดไอโอดีนเป็นการละเมิดการก่อตัวของฮอร์โมนต่อมไทรอยด์และการลดลงของความเข้มของการเผาผลาญ การลดการไหลของฟลูออรีนด้วยน้ำและอาหารในระดับใหญ่กำหนดการละเมิดการต่ออายุของเคลือบฟันของฟันและเป็นผลให้ความบกพร่องในการฟันผุ ตะกั่วเป็นพิษเกือบสำหรับสิ่งมีชีวิตทั้งหมด สาเหตุที่เกินความเสียหายต่อสมองและระบบประสาทส่วนกลางซึ่งแสดงถึงการสูญเสียวิสัยทัศน์และการได้ยินนอนไม่หลับภาวะไตวายชักและยังสามารถนำไปสู่โรคอัมพาตและโรคนี้เป็นโรคมะเร็ง ตะกั่วผู้บุกเบิกเฉียบพลันจะมาพร้อมกับภาพหลอนฉับพลันและจบลงด้วยอาการโคม่าและความตาย
การขาดแมโครและไมโครไลน์ย์สามารถชดเชยได้โดยการเพิ่มเนื้อหาในอาหารและน้ำดื่มรวมถึงการรับ การเตรียมยา. ดังนั้นไอโอดีนมีอยู่ในอาหารทะเลและเกลือไอโอดีนแคลเซียม - ในเปลือกไข่ ฯลฯ
ความสัมพันธ์ระหว่างโครงสร้างและหน้าที่ของสารอนินทรีย์และสารอินทรีย์ (โปรตีน, กรดนิวคลีอิก, คาร์โบไฮเดรต, ไขมัน lipids) ซึ่งรวมอยู่ในเซลล์ บทบาทของสารเคมีในเซลล์และร่างกายมนุษย์
สารอนินทรีย์
องค์ประกอบของเซลล์เคมีก่อให้เกิดสารประกอบที่แตกต่างกัน - อนินทรีย์และอินทรีย์ สารของเซลล์อนินทรีย์รวมถึงน้ำเกลือแร่กรด ฯลฯ และเป็นอินทรีย์ - โปรตีน, กรดนิวคลีอิก, คาร์โบไฮเดรต, ไขมัน, atp, วิตามินและอื่น ๆ
น้ำ (H 2 O) เป็นสารอนินทรีย์ที่พบมากที่สุดที่มีคุณสมบัติทางเคมีกายภาพที่เป็นเอกลักษณ์ เธอไม่มีรสนิยมหรือสีไม่มีกลิ่น ความหนาแน่นและความหนืดของสารทั้งหมดมีการประมาณด้วยน้ำ เช่นเดียวกับสารอื่น ๆ น้ำสามารถอยู่ในสามรัฐรวม: ของแข็ง (น้ำแข็ง), ของเหลวและก๊าซ (คู่) จุดหลอมเหลวคือ $ 0 ° $ C จุดเดือดคือ $ 100 ° C อย่างไรก็ตามการละลายในน้ำของสารอื่น ๆ สามารถเปลี่ยนลักษณะเหล่านี้ได้ ความจุความร้อนน้ำยังมีขนาดใหญ่พอ - 4200 kJ / mol · k ซึ่งให้โอกาสเธอในการมีส่วนร่วมในกระบวนการเทอร์โมนัล ในโมเลกุลของน้ำอะตอมไฮโดรเจนตั้งอยู่ที่มุมของ $ 105 ° $ ในขณะที่คู่อิเล็กทรอนิกส์ทั่วไปล่าช้าด้วยอะตอมออกซิเจนอออกได้มากขึ้น สิ่งนี้ทำให้คุณสมบัติของไดโพลของโมเลกุลน้ำ (หนึ่งในจุดสิ้นสุดของพวกเขาถูกเรียกเก็บในเชิงบวกและอื่น ๆ เป็นลบ) และความเป็นไปได้ของการก่อตัวระหว่างโมเลกุลของน้ำของพันธะไฮโดรเจน กริปของโมเลกุลของน้ำที่เหมาะกับปรากฏการณ์ของความตึงเครียดของพื้นผิว, ภูมิทัศน์เส้นเลือดฝอยและน้ำเป็นตัวทำละลายสากล เป็นผลให้สารทั้งหมดแบ่งออกเป็นน้ำที่ละลายน้ำได้ (hydrophilic) และไม่ละลายในมัน (ไม่ชอบน้ำ) ต้องขอบคุณคุณสมบัติที่เป็นเอกลักษณ์เหล่านี้จะกำหนดไว้ล่วงหน้าว่าน้ำกลายเป็นพื้นฐานของชีวิตบนโลก

ปริมาณน้ำเฉลี่ยในเซลล์ของร่างกายไม่เหมือนกันและสามารถเปลี่ยนแปลงได้ตามอายุ ดังนั้นในตัวอ่อนของมนุษย์สองเดือนปริมาณน้ำในเซลล์ถึง 97.5% ในแปดเดือน - 83% ทารกแรกเกิดลดลงเป็น 74% และเฉลี่ย 66% เป็นค่าเฉลี่ย 66% ใน ผู้ใหญ่ อย่างไรก็ตามเซลล์ของร่างกายแตกต่างกันในปริมาณน้ำ ดังนั้นกระดูกมีประมาณ 20% ของน้ำในตับ - 70% และในสมอง - 86% โดยทั่วไปเราสามารถพูดได้ว่า ความเข้มข้นของน้ำในเซลล์เป็นสัดส่วนโดยตรงกับความเข้มของการเผาผลาญ.
เกลือแร่ อาจอยู่ในสถานะที่ละลายหรือไม่มีปัญหา เกลือที่ละลายน้ำได้ แยกจากกันกับไอออน - ไอออนบวกและไอออนไอออน ไอออนบวกที่สำคัญที่สุดคือโพแทสเซียมและโซเดียมไอออนอำนวยความสะดวกในการถ่ายโอนสารผ่านเมมเบรนและมีส่วนร่วมในการเกิดขึ้นและดำเนินการกระตุ้นประสาท เช่นเดียวกับแคลเซียมไอออนซึ่งมีส่วนร่วมในกระบวนการลดเส้นใยกล้ามเนื้อและการแข็งตัวของเลือด แมกนีเซียมรวมอยู่ในคลอโรฟิลล์; เหล็กซึ่งเป็นส่วนหนึ่งของชุดของโปรตีนรวมถึงฮีโมโกลบิน แอนไอออนที่สำคัญที่สุดคือประจุลบฟอสเฟตซึ่งเป็นส่วนหนึ่งของกรด ATP และนิวเคลียสและสารตกค้างของกรดเคลือบเงาซึ่งทำให้การแกว่งของค่า pH ของปานกลาง ไอออนของเกลือแร่ให้และเจาะน้ำในกรงและมันอยู่ในนั้น หากในสื่อความเข้มข้นของเกลือต่ำกว่าในเซลล์แล้วน้ำจะแทรกซึมเข้าไปในเซลล์ นอกจากนี้ไอออนจะถูกกำหนดโดยสมบัติของบัฟเฟอร์ของไซโตพลาสซึมนั่นคือความสามารถในการรักษาความมั่นคงของค่า pH อัลคาไลน์ที่อ่อนแอของไซโตพลาสซึมแม้จะมีการก่อตัวอย่างต่อเนื่องในผลิตภัณฑ์กรดและอัลคาไลน์
เกลือที่ไม่ละลายน้ำ (Caco 3, CA 3 (PO 4) 2 ฯลฯ ) รวมอยู่ในกระดูกฟันเปลือกหอยและเปลือกหอยของสัตว์ยูนิแคร์และหลายเซลล์
นอกจากนี้สารประกอบอนินทรีย์อื่น ๆ เช่นกรดและออกไซด์สามารถผลิตได้ในสิ่งมีชีวิต ดังนั้น Chinck ของเซลล์กระเพาะอาหารของมนุษย์ผลิตกรดไฮโดรคลอริกซึ่งเปิดใช้งานเอนไซม์ย่อยอาหารของ Pepsin และ Silicon Oxide สร้างความประทับใจให้กับผนังเซลล์ของหางม้าและรูปแบบเปลือกคลื่นไดอะตอม ใน ปีที่แล้ว บทบาทของไนโตรเจนออกไซด์ (II) ยังถูกตรวจสอบในการส่งสัญญาณในเซลล์และร่างกาย
สารอินทรีย์
ลักษณะโดยรวมของเซลล์สารอินทรีย์
เซลล์อินทรีย์ของเซลล์สามารถแสดงเป็นโมเลกุลที่ค่อนข้างง่ายและซับซ้อนมากขึ้น ในกรณีที่โมเลกุลที่ซับซ้อน (Macromolecule) เกิดขึ้นจากโมเลกุลที่ง่ายกว่าซ้ำหลายครั้งเรียกว่า โพลิเมอร์และหน่วยโครงสร้าง - โมโนเมอร์. ขึ้นอยู่กับว่าจะทำซ้ำหรือไม่มีลิงค์ของโพลิเมอร์พวกเขาจะถูกอ้างถึง ปกติ หรือ ผิดปกติ. โพลิเมอร์สูงถึง 90% ของมวลของเซลล์แห้ง พวกเขาเกี่ยวข้องกับสามคลาสหลักของสารประกอบอินทรีย์ - คาร์โบไฮเดรต (polysaccharides), โปรตีนและกรดนิวคลีอิก โพลีเมอร์ปกติเป็นโพลีแซคคาไรด์และโปรตีนและกรดนิวคลีอิก - ผิดปกติ ในโปรตีนและกรดนิวคลีอิกลำดับโมโนเมอร์มีความสำคัญอย่างยิ่งเนื่องจากทำหน้าที่ให้ข้อมูล

คาร์โบไฮเดรต
คาร์โบไฮเดรต - เหล่านี้เป็นสารประกอบอินทรีย์ซึ่งรวมถึงองค์ประกอบทางเคมีสามชิ้น - คาร์บอนไฮโดรเจนและออกซิเจนแม้ว่าคาร์โบไฮเดรตจำนวนหนึ่งยังมีไนโตรเจนหรือซัลเฟอร์ สูตรคาร์โบไฮเดรตทั้งหมด - กับ m (h 2 o) n. พวกเขาแบ่งออกเป็นคาร์โบไฮเดรตที่เรียบง่ายและซับซ้อน
คาร์โบไฮเดรตง่าย ๆ (monosaccharides) มีโมเลกุลน้ำตาลเดียวที่เป็นไปไม่ได้ที่จะแยกออกเป็นเกมที่ง่ายกว่า เหล่านี้เป็นสารผลึกรสชาติหวานและละลายได้ดีในน้ำ monosaccharides ใช้ส่วนที่ใช้งานอยู่ในการแลกเปลี่ยนสารในเซลล์และเป็นส่วนหนึ่งของคาร์โบไฮเดรตที่ซับซ้อน - oligosaccharides และ polysaccharides
Monosaccharides จำแนกตามจำนวนอะตอมคาร์บอน (C 3 -C 9) ตัวอย่างเช่น ทำให้เพนโตส (จาก 5) และ hxosis (จาก 6) Penosas รวมถึง Roose และ Deoxyribosis ย่าง มันเป็นส่วนหนึ่งของ RNA และ ATP Deoxyribese มันเป็นองค์ประกอบของ DNA hxosis (จาก 6 ชั่วโมง 12 o 6) เป็นกลูโคสฟรักโทสกาแลคโตส ฯลฯ กลูโคส (น้ำตาลองุ่น) พบได้ในทุกสิ่งมีชีวิตรวมถึงในเลือดมนุษย์เนื่องจากเป็นพลังงานสำรอง มันเป็นส่วนหนึ่งของน้ำตาลที่ซับซ้อนมากมาย: ซูโครส, แลคโตส, มอลโตส, แป้ง, เซลลูโลส, ฯลฯ ฟรักโทส(น้ำตาลผลไม้) ในความเข้มข้นที่ยิ่งใหญ่ที่สุดมีอยู่ในผลไม้น้ำผึ้งหัวผักกาดน้ำตาลรูท ไม่เพียง แต่ใช้ส่วนที่ใช้งานในกระบวนการเผาผลาญ แต่ยังเป็นส่วนหนึ่งของซูโครสและโพลีแซคคาไรด์บางส่วนเช่นอินซูลิน
monosaccharides ส่วนใหญ่สามารถทำปฏิกิริยากับกระจกสีเงินและคืนค่าทองแดงเมื่อมีการเพิ่มของเหลวด้วย (ส่วนผสมของทองแดง (ii) ซัลเฟตและโซเดียมโพแทสเซียมโซเดียม) และเดือด

ถึง oligosaccharidam เชื่อว่าคาร์โบไฮเดรตที่เกิดจากสารตกค้างหลายชนิดของ monosaccharides พวกเขาส่วนใหญ่ละลายได้ดีในน้ำและรสชาติขนมหวาน ขึ้นอยู่กับจำนวนของสารตกค้างเหล่านี้ disaccharides มีความโดดเด่น (สองสารตกค้าง), trisaccharides (สาม) และอื่น ๆ disaccharide รวมถึงซูโครส, แลคโตส, มอลโตส ฯลฯ ซากศพ(น้ำตาลบีทหรือน้ำตาล) ประกอบด้วยกลูโคสและฟรุกโตสที่ตกค้างพบในอวัยวะพื้นฐานของพืชบางชนิด โดยเฉพาะอย่างยิ่งซูโครสหลายตัวในรากของหัวผักกาดน้ำตาลและอ้อยจากที่พวกเขาได้รับจากวิธีอุตสาหกรรม มันทำหน้าที่เป็นมาตรฐานของความหวานของคาร์โบไฮเดรต แลคโตส, หรือ น้ำตาลนมซึ่งเกิดจากเศษซากกลูโคสและกาแลคโตสมีอยู่ในนมแม่และนมวัว มอลโตส (น้ำตาลมอลต์) ประกอบด้วยสารตกค้างของกลูโคสสองตัว มันเกิดขึ้นในกระบวนการของการแยกโพลีแซคคาไรด์ในเมล็ดพืชและในระบบย่อยอาหารของมนุษย์ใช้ในการผลิตเบียร์

polysaccharides - สิ่งเหล่านี้เป็น biopolymers ที่โมโนเมอร์เป็นส่วนที่เหลือของ mono- หรือ disaccharides โพลีแซคคาไรด์ส่วนใหญ่ไม่ละลายในน้ำและด้อยโอกาส เหล่านี้รวมถึงแป้งไกลโคเจนเซลลูโลสและไคติน แป้ง - นี่คือสารผงสีขาวที่ไม่เปียกน้ำ แต่การระงับน้ำร้อนขึ้นรูปน้ำร้อน ในความเป็นจริงแล้วแป้งประกอบด้วยโพลิเมอร์สองตัว - อะมิลีสที่แตกสลายน้อยกว่าและอะไมโลปินสาขามากขึ้น (รูปที่ 2.9) โมโนเมอร์เช่น Amylose และ Amylopectin เป็นกลูโคส แป้งเป็นอาหารสำรองหลักของพืชซึ่งในจำนวนมากสะสมในเมล็ดผลไม้, หัว, เหง้าและพืชอื่น ๆ ปฏิกิริยาที่มีคุณภาพสูงต่อแป้งเป็นปฏิกิริยากับไอโอดีนที่แป้งถูกทาสีในสีฟ้า - ม่วง

ไกลโคเจน (แป้งสัตว์) เป็นโพลีแซคคาไรด์สำรองของสัตว์และเห็ดซึ่งในปริมาณที่มากที่สุดสะสมในกล้ามเนื้อและตับ นอกจากนี้ยังไม่ละลายในน้ำและรสชาติ Glycogen Monomer เป็นกลูโคส เมื่อเทียบกับโมเลกุลแป้งโมเลกุลไกลโคเจนนั้นมีกิ่งมากขึ้น
เซลลูโลส, หรือ เซลลูโลส- การสนับสนุนหลัก polysaccharide ของพืช โมโนเมอร์เซลลูโลสเป็นกลูโคส โมเลกุลเซลลูโลสในรูปแบบการรวมกลุ่มซึ่งรวมอยู่ในผนังเซลล์ของพืช เซลลูโลสเป็นพื้นฐานของไม้มันถูกใช้ในการก่อสร้างในการผลิตเนื้อเยื่อกระดาษแอลกอฮอล์และสารอินทรีย์จำนวนมาก เซลลูโลสมีความเฉื่อยชาทางเคมีและไม่ละลายในกรดหรืออัลคาลิส นอกจากนี้ยังไม่ได้แยกเอนไซม์ของระบบย่อยอาหารของมนุษย์ แต่แบคทีเรียของลำไส้ใหญ่มีส่วนร่วมในการย่อยอาหาร นอกจากนี้ไฟเบอร์ช่วยกระตุ้นการลดการลดลงของผนังของระบบทางเดินอาหารให้การสนับสนุนการปรับปรุงงานของมัน

ไคติน - นี่คือ polysaccharide ที่โมโนเมอร์เป็น monosaccharide ที่มีไนโตรเจน มันเป็นส่วนหนึ่งของผนังเซลล์ของเห็ดและสัตว์ผ่าตัดที่อ่อนโยน ในระบบย่อยอาหารของมนุษย์เอนไซม์การย่อยอาหารของไคตินก็หายไปก็มีเพียงแบคทีเรียบางชนิดเท่านั้น
ฟังก์ชั่นของคาร์โบไฮเดรต คาร์โบไฮเดรตดำเนินการในพลาสติก (การก่อสร้าง), พลังงาน, ถุงน่องและฟังก์ชั่นอ้างอิง พวกเขาสร้างผนังเซลล์ของพืชและเห็ด ค่าพลังงานของการแยก 1 กรัมคาร์โบไฮเดรตคือ 17.2 kJ กลูโคสฟรุกโตสซูโครสแป้งและไกลโคเจนเป็นสัดส่วนอะไหล่ คาร์โบไฮเดรตยังสามารถเป็นส่วนหนึ่งของไขมันที่ซับซ้อนและโปรตีนสร้าง glycolipids และ glycoproteins โดยเฉพาะในเยื่อหุ้มเซลล์ ไม่มีอะไรสำคัญน้อยกว่าคือบทบาทของคาร์โบไฮเดรตในการรับรู้ระหว่างเซลล์และการรับรู้สัญญาณของสภาพแวดล้อมภายนอกเนื่องจากอยู่ในองค์ประกอบของ Glycoproteins ดำเนินการฟังก์ชั่นตัวรับ
lipids
lipids - นี่เป็นความแตกต่างในอัตราส่วนทางเคมีของสารน้ำหนักโมเลกุลต่ำที่มีคุณสมบัติที่ไม่ชอบน้ำ สารเหล่านี้ไม่ละลายในน้ำเป็นอิมัลชันในนั้น แต่ในเวลาเดียวกันพวกเขาก็ละลายได้ดีในตัวทำละลายอินทรีย์ น้ำมันไขมันในการสัมผัสพวกเขาหลายคนทิ้งร่องรอยที่ไม่แห้ง พร้อมกับโปรตีนและคาร์โบไฮเดรตพวกเขาเป็นหนึ่งในองค์ประกอบหลักของเซลล์ เนื้อหาของไขมันในเซลล์ต่าง ๆ ไม่เหมือนกันโดยเฉพาะอย่างยิ่งพวกเขาหลายคนในเมล็ดและผลไม้ของพืชบางชนิดในตับหัวใจเลือด
ขึ้นอยู่กับโครงสร้างของโมเลกุลไขมันพวกเขาแบ่งออกเป็นเรียบง่ายและซับซ้อน ถึง ง่าย LIPID รวมถึงไขมันที่เป็นกลาง (ไขมัน), แว็กซ์และสเตียรอยด์ มีความซับซ้อน LIPIDS มีส่วนประกอบ Nelipid อื่น ที่สำคัญที่สุดของสิ่งเหล่านี้คือฟอสโฟไลปิด, glycolipids ฯลฯ
อ้วน. นี่คือเอสเทอร์ของกลีเซอรี่แอลกอฮอล์ Trochaty และกรดไขมันที่สูงขึ้น กรดไขมันส่วนใหญ่มีอะตอม 14-22 คาร์บอน ในหมู่พวกเขาทั้งที่อุดมไปด้วยและไม่อิ่มตัวนั่นคือมีพันธะคู่ จากกรดไขมันอิ่มตัวส่วนใหญ่พบฝ่ามือและสเตียโนวยาและจากไม่อิ่มตัว - โอเลอิค กรดไขมันไม่อิ่มตัวบางชนิดไม่ได้สังเคราะห์ในร่างกายมนุษย์หรือสังเคราะห์ในปริมาณที่ไม่เพียงพอดังนั้นจึงขาดไม่ได้ เศษของกลีเซอรีนก่อตัวเป็น "หัว" ที่ชอบน้ำและซากกรดไขมันนั้นเป็น "หาง" ที่ไม่ชอบน้ำ

ไขมันจะดำเนินการในเซลล์ส่วนใหญ่ในการทำงานของสต็อกและทำหน้าที่เป็นแหล่งพลังงาน พวกเขาอุดมไปด้วยเส้นใยไขมันใต้ผิวหนังการทำค่าเสื่อมราคาและฟังก์ชั่นฉนวนความร้อนและในสัตว์น้ำ - ยังเพิ่มการลอยตัว Pynes ของพืชส่วนใหญ่มีกรดไขมันไม่อิ่มตัวเป็นผลมาจากสภาพที่เป็นของเหลวและเรียกว่า น้ำมัน. น้ำมันมีอยู่ในเมล็ดพืชจำนวนมากเช่นดอกทานตะวันถั่วเหลืองข่มขืน ฯลฯ
ขี้ผึ้ง - นี่คือเอสเทอร์และส่วนผสมของกรดไขมันและแอลกอฮอล์ไขมัน ในพืชพวกเขาสร้างภาพยนตร์บนพื้นผิวของแผ่นงานซึ่งป้องกันการระเหยการรุกของเชื้อโรค ฯลฯ ในสัตว์จำนวนมากพวกเขาครอบคลุมร่างกายหรือทำหน้าที่สร้างเซลล์
ถึง สเตียรอยด์ ไขมันเหล่านี้เช่นคอเลสเตอรอลเป็นองค์ประกอบที่บังคับของเยื่อหุ้มเซลล์เช่นเดียวกับฮอร์โมนเพศ Estradiol, ฮอร์โมนเพศชาย, วิตามินดี, ฯลฯ
Phospholipidsนอกเหนือไปจากการตกค้างของกลีเซอรอลและกรดไขมันมีสารตกค้างของกรดออร์โธฟอสฟอริก พวกเขาเป็นส่วนหนึ่งของเยื่อหุ้มเซลล์และให้คุณสมบัติอุปสรรคของพวกเขา
glycolipids นอกจากนี้ยังเป็นองค์ประกอบของเมมเบรน แต่เนื้อหาของพวกเขามีขนาดเล็กที่นั่น ส่วน nelipid ของ glycolipids คือคาร์โบไฮเดรต
ฟังก์ชั่นไขมัน ไขมันจะดำเนินการในพลาสติก (ก่อสร้าง), พลังงาน, ถุงเท้ายาว, ป้องกัน, ขับถ่ายและการกำกับดูแลนอกจากนี้พวกเขาเป็นวิตามิน นี่เป็นองค์ประกอบที่จำเป็นของเยื่อหุ้มเซลล์ เมื่อแยกไขมัน 1 กรัมพลังงาน 38.9 kj ของพลังงานถูกไฮไลต์ พวกเขาวางในสต็อกในอวัยวะต่าง ๆ ของพืชและสัตว์ นอกจากนี้เส้นใยไขมันใต้ผิวหนังช่วยปกป้องอวัยวะภายในจาก Supercooling หรือความร้อนสูงเกินไปเช่นเดียวกับแรงกระแทก ฟังก์ชั่นการกำกับดูแลไขมันเกี่ยวข้องกับความจริงที่ว่าบางคนเป็นฮอร์โมน ร่างกายไขมันแมลงทำหน้าที่เพื่อเน้น
โปรตีน
โปรตีน - เหล่านี้เป็นสารประกอบโมเลกุลสูง Biopolymers ที่โมโนเมอร์เป็นกรดอะมิโนที่เกี่ยวข้องกับพันธะเปปไทด์
กรดอะมิโน โทรหาสารประกอบอินทรีย์ที่มีกลุ่มอะมิโนกลุ่มคาร์บอกซิลและหัวรุนแรง รวมประมาณ 200 กรดอะมิโนพบในธรรมชาติซึ่งแตกต่างกันในอนุมูลและ ตำแหน่งซึ่งกันและกัน กลุ่มการทำงาน แต่มีเพียง 20 คนเท่านั้นที่อาจเป็นส่วนหนึ่งของโปรตีน กรดอะมิโนดังกล่าวเรียกว่า โปรตีน

น่าเสียดายที่กรดอะมิโนโปรตีนทั้งหมดไม่สามารถสังเคราะห์ในร่างกายมนุษย์ดังนั้นพวกเขาจึงแบ่งออกเป็นเปลี่ยนได้และขาดไม่ได้ กรดอะมิโนที่ถอดเปลี่ยนได้ เกิดขึ้นในร่างกายมนุษย์ในปริมาณที่ต้องการและ ขาดไม่ได้ - ไม่ใช่. พวกเขาจะต้องมาพร้อมกับอาหาร แต่สามารถสังเคราะห์จุลินทรีย์ในลำไส้ได้บางส่วน กรดอะมิโนที่ขาดไม่ได้อย่างเต็มที่หมายเลข 8 เหล่านี้รวมถึงการมัลติ, Isolecin, Leucine, Lizin, Methionine, Threonine, Tryptophan และ Phenylalanine แม้จะมีความจริงที่ว่ากรดอะมิโนโปรตีนทั้งหมดจะถูกสังเคราะห์ในพืชโปรตีนผักมีข้อบกพร่องเนื่องจากพวกเขาไม่มีกรดอะมิโนที่สมบูรณ์นอกเหนือจากการปรากฏตัวของโปรตีนในชิ้นส่วนพืชของพืชไม่ค่อยเกิน 1-2% ของ มวล ดังนั้นจึงจำเป็นต้องกินโปรตีนไม่เพียง แต่ผัก แต่ยังมาจากสัตว์
ลำดับของกรดอะมิโนสองตัวที่เกี่ยวข้องกับพันธะเปปไทด์เรียกว่า ญาติ, จากสาม - ตริหมาส ฯลฯ ในบรรดาเปปไทด์มีสารประกอบสำคัญเช่นฮอร์โมน (oxytocin, vasopressin), ยาปฏิชีวนะ, ฯลฯ โซ่ของกรดอะมิโนมากกว่ายี่สิบอะมิโน โพลีเปปไทด์และโพลีเปปไทด์ที่มีสารตกค้างของกรดอะมิโนมากกว่า 60 ชนิดเป็นโปรตีน

ระดับขององค์กรโครงสร้างของโปรตีน โปรตีนอาจมีโครงสร้างประถมศึกษาระดับอุดมศึกษาระดับอุดมศึกษาระดับอุดมศึกษาและ Quaternary
โครงสร้างโปรตีนหลัก - นี่คือ ลำดับเชิงเส้นของกรดอะมิโนเชื่อมต่อด้วยพันธะเปปไทด์ ในที่สุดโครงสร้างหลักจะกำหนดความจำเพาะของโปรตีนและเอกลักษณ์เพราะแม้ว่าเราจะคิดว่าในโปรตีนขนาดกลางมีสารตกค้างของกรดอะมิโน 500 รายการจำนวนชุดค่าผสมที่เป็นไปได้คือ 20 500 ดังนั้นการเปลี่ยนแปลงในตำแหน่งของที่ อย่างน้อยหนึ่งอะมิโนกรดในโครงสร้างหลักจะมีการเปลี่ยนแปลงในโครงสร้างรองและที่สูงกว่าเช่นเดียวกับคุณสมบัติของโปรตีนโดยรวม

คุณสมบัติของโครงสร้างของโปรตีนกำหนดการวางเชิงพื้นที่ - การเกิดขึ้นของโครงสร้างทุติยภูมิและตติยภูมิ
โครงสร้างรอง เป็นการวางอวกาศของโมเลกุลโปรตีนเช่น เกลียว หรือ พับถือโดยพันธะไฮโดรเจนระหว่างออกซิเจนและอะตอมไฮโดรเจนของกลุ่มเปปไทด์ของแหลมหรือเท่าที่แตกต่างกัน โปรตีนจำนวนมากมีส่วนยาวมากหรือน้อยกับโครงสร้างรอง นี่คือตัวอย่างเช่น keratines ของผมและเล็บ, ผ้าไหมไฟโบริน
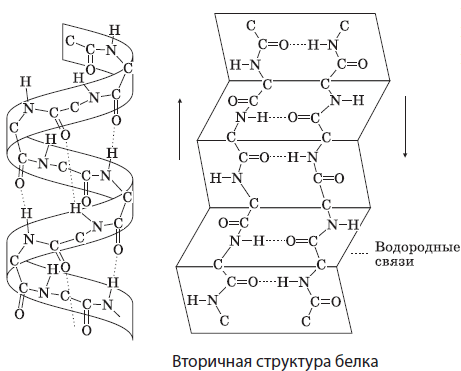
โครงสร้างตติยภูมิ กระรอก ( ลูกโลกนอกจากนี้ยังเป็นรูปแบบของการวางเชิงพื้นที่ของโซ่โพลีเปปไทด์ที่ถือโดย hydrophobic, ไฮโดรเจน, ซัลไฟด์ (S) และการเชื่อมต่ออื่น ๆ มันเป็นลักษณะของโปรตีนสิ่งมีชีวิตส่วนใหญ่เช่นกล้ามเนื้อ Moglobin

โครงสร้าง Quaternary - ที่ซับซ้อนที่สุดที่เกิดขึ้นจากโซ่ polypeptide หลายแห่งนั้นประกอบไปด้วยพันธบัตรเดียวกันกับที่อยู่ในระดับอุดมศึกษา (ไม่ชอบน้ำ, ไอออนไอออนและไฮโดรเจน) รวมถึงการโต้ตอบที่อ่อนแออื่น ๆ โครงสร้าง Quaternary เป็นลักษณะของโปรตีนบางอย่างเช่นฮีโมโกลบินคลอโรฟิล ฯลฯ

ในรูปแบบของโมเลกุลแยกแยะความแตกต่าง เกี่ยวกับฟริลลารี และ เป็นทรงกลม โปรตีน คนแรกของพวกเขามีความยาวเช่นเนื้อเยื่อเกี่ยวพันคอลลาเจนหรือ keratines ของผมและเล็บ โปรตีนทรงกลมมีรูปร่างที่ยุ่งเหยิง (ลูกโลก) เช่นกล้ามเนื้อ myoglobin
โปรตีนที่เรียบง่ายและซับซ้อน กระรอกสามารถ ง่าย และ ซับซ้อน โปรตีนง่าย ๆ ประกอบด้วยกรดอะมิโนเพียงอย่างเดียวในขณะที่ มีความซับซ้อน โปรตีน (ไลโปโปรตีน, chromoproteins, glycoproteins, nucleoproteins, ฯลฯ ) มีส่วนโปรตีนและไม่เป็นที่ไม่มีวิธี chromoprotein มีส่วนที่ไม่ได้ทาสีที่ทาสี เหล่านี้รวมถึง hemoglobin, mioglobin, คลอโรฟิล, ไซโตโครม, เป็นส่วนหนึ่งของฮีโมโกลบิน, โซ่โพลีเปปไทด์สี่ของโปรตีนของ Globin มีความเกี่ยวข้องกับส่วนที่ไม่ได้ค้นพบ - กับชายเสื้อที่อยู่ตรงกลางซึ่งเป็นไอออนเหล็ก ที่ให้ฮีโมโกลบินเป็นสีแดง ส่วนที่ไม่ถูกค้นพบ ไลโปโปรตีน lipid และ glycoproteins - คาร์โบไฮเดรต ไลโปโปรตีนและ glycoproteins รวมอยู่ในเยื่อหุ้มเซลล์ นิวเคลียส พวกเขามีความซับซ้อนของโปรตีนและกรดนิวคลีอิก (DNA และ RNA) พวกเขาทำหน้าที่ที่จำเป็นในการจัดเก็บและส่งข้อมูลและการส่งข้อมูลทางพันธุกรรม
คุณสมบัติของโปรตีน โปรตีนหลายชนิดละลายได้ดีในน้ำ แต่มีอยู่ในหมู่พวกเขาและผู้ที่ละลายในการแก้ปัญหาของเกลือ, ด่าง, กรดหรือตัวทำละลายอินทรีย์ โครงสร้างของโมเลกุลโปรตีนและกิจกรรมการทำงานของมันขึ้นอยู่กับสภาพแวดล้อม การสูญเสียโมเลกุลโปรตีนของโครงสร้างในขณะที่รักษาหลักเรียกว่า การเสียสละ.

การสูญเสียเกิดขึ้นเนื่องจากการเปลี่ยนแปลงของอุณหภูมิ, ค่า pH, ความดันบรรยากาศภายใต้อิทธิพลของกรด, ด่าง, เกลือของโลหะหนัก, ตัวทำละลายอินทรีย์ ฯลฯ กระบวนการย้อนกลับของการฟื้นฟูโครงสร้างรองและที่สูงกว่าเรียกว่า renaturaอย่างไรก็ตามมันเป็นไปไม่ได้เสมอไป การทำลายล้างอย่างเต็มรูปแบบของโมเลกุลโปรตีนเรียกว่า การทำลาย.
ฟังก์ชั่นโปรตีนโปรตีนจะดำเนินการในเซลล์จำนวนหนึ่งของฟังก์ชั่น: พลาสติก (ก่อสร้าง), ตัวเร่งปฏิกิริยา (เอนไซม์), พลังงาน, การส่งสัญญาณ (รับ), หดตัว (มอเตอร์), การขนส่ง, ป้องกัน, กฎระเบียบและการเก็บรักษา
ฟังก์ชั่นการก่อสร้างของโปรตีนมีความเกี่ยวข้องกับการปรากฏตัวของพวกเขาในเยื่อหุ้มเซลล์และองค์ประกอบโครงสร้างของเซลล์ พลังงาน - เกิดจากความจริงที่ว่าเมื่อแยกโปรตีน 1 กรัมเปิดตัว 17.2 kJ ของพลังงาน โปรตีนตัวรับเมมเบรนมีส่วนร่วมในการรับรู้สัญญาณด้านสิ่งแวดล้อมและการส่งผ่านของเซลล์เช่นเดียวกับการจดจำระหว่างเซลล์ หากไม่มีโปรตีนการเคลื่อนไหวของเซลล์และสิ่งมีชีวิตโดยทั่วไปเป็นไปไม่ได้เนื่องจากเป็นพื้นฐานของ Flagella และ Cilia และยังให้การหดตัวของกล้ามเนื้อและการเคลื่อนไหวของส่วนประกอบของเซลล์ ในเลือดของมนุษย์และสัตว์จำนวนมากฮีโมโกลบินโอนออกซิเจนและเป็นส่วนหนึ่งของก๊าซคาร์บอนไดออกไซด์, โปรตีนอื่น ๆ ไอออนและอิเล็กตรอน บทบาทการป้องกันของโปรตีนที่เชื่อมต่อกันก่อนอื่นด้วยภูมิคุ้มกันเนื่องจากโปรตีน Interferon มีความสามารถในการทำลายไวรัสจำนวนมากและแอนติบอดีโปรตีนที่ปราบปรามการพัฒนาของแบคทีเรียและตัวแทนต่างด้าวอื่น ๆ ในบรรดาโปรตีนและเปปไทด์ฮอร์โมนจำนวนมากเช่นฮอร์โมนของตับอ่อน - อินซูลินควบคุมความเข้มข้นของกลูโคสในเลือด ในบางสิ่งมีชีวิตโปรตีนสามารถเลื่อนออกไปในการสำรองเช่นถั่วในเมล็ดหรือโปรตีนไข่ไก่
กรดนิวคลีอิก
กรดนิวคลีอิก- สิ่งเหล่านี้เป็น Biopolymers ที่โมโนเมอร์เป็นนิวคลีโอไทด์ ปัจจุบันมีกรดนิวคลีอิกสองประเภท: Ribonucleic (RNA) และ Deoxyribonuclear (DNA)
นิวคลีโอไทด์ เกิดจากฐานไนโตรเจนที่ตกค้างของการเพนุจน้ำตาลและสารตกค้างของกรดออร์โธฟอสฟอริก คุณสมบัติของนิวคลีโอไทด์ส่วนใหญ่จะถูกกำหนดโดยฐานไนโตรเจนที่รวมอยู่ในองค์ประกอบของพวกเขาดังนั้นแม้แต่นิวคลีโอไทด์ตามเงื่อนไขจะถูกระบุโดยตัวอักษรตัวแรกของชื่อ องค์ประกอบของนิวคลีโอไทด์อาจรวมถึงฐานไนโตรเจนห้าแห่ง: Adenine (A), Guanine (G), Timin (t), uracil (Y) และ Cytosin (c) Pentoses of Nucleotides - Ribosis และ deoxyribosis - กำหนดว่านิวคลีโอไทด์ใดเป็น ribonucleotide หรือ deoxyribonucleotide Ribonucleotides เป็น RNA Monomers สามารถทำหน้าที่เป็นโมเลกุลสัญญาณ (CAMF) และเพื่อเข้าสู่สารประกอบ macroergic เช่น ATP และ Coenzymes เช่น Nadf, OB, FAD, ฯลฯ และ Deoxyribonucleotides เป็นส่วนหนึ่งของ DNA

กรด Deoxyribonucleic (DNA) - Biopolymer สองที่ติดอยู่ซึ่งโมโนเมอร์เป็น deoxyribonucleotides องค์ประกอบของ deoxyribonucleotides ประกอบด้วยเพียงสี่ฐานไนโตรเจนที่เป็นไปได้ - Adenine (A), Timin (t), Guanine (g) หรือ cytosine (c) เช่นเดียวกับสารตกค้าง deoxyribose และกรดออร์โปลิสโฟโต นิวคลีโอไทด์ในวงจร DNA เชื่อมต่อกันผ่านซากของกรดออร์โธฟอสฟอริกการสร้างการสื่อสารของฟอสเฟตสเตอร์ ในการก่อตัวของโมเลกุลสองที่ติดอยู่ฐานไนโตรเจนถูกนำเข้าสู่โมเลกุล อย่างไรก็ตามการปลดปล่อยของวงจร DNA ที่เกิดขึ้นตามฐานของวงจรไนโตรเจนแบบสุ่มรวมกันโดยหลักการของการเสริม: Adenine เชื่อมต่อกับ Thymine กับสองพันธะไฮโดรเจน (A \u003d T) และ Guanine ที่มี Cytosine เป็นสาม (g $ ≡ $ c)

สำหรับเธอถูกติดตั้ง กฎของ Chargaff:
- จำนวน Nucleotides DNA ที่มี adenine เท่ากับปริมาณนิวคลีโอไทด์ที่มีโทจิน (A \u003d t)
- จำนวนของ Nucleotides DNA ที่มี Guanine เท่ากับจำนวนนิวคลีโอไทด์ที่มี cytosine (R $ ≤ $ c)
- ผลรวมของ deoxyribonucleotides ที่มี adenine และ guanine เท่ากับจำนวน deoxyribonucleotides ที่มี thymine และ cytosin (a + g \u003d t + c)
- อัตราส่วนของปริมาณ deoxyribonucleotides ที่มี adenine และ timin กับผลรวมของ deoxyribonucleotides ที่มี guanine และ cytosin ขึ้นอยู่กับประเภทของสิ่งมีชีวิต
โครงสร้าง DNA ถูกถอดรหัสโดย F. Creek และ D. Watson (รางวัลโนเบลสาขาสรีรวิทยาและยา 2505) ตามรุ่นของพวกเขา Molecule DNA เป็น Human Helix คู่ ระยะห่างระหว่างนิวคลีโอไทด์ในวงจร DNA คือ 0.34 nm

คุณสมบัติที่สำคัญที่สุดของ DNA คือความสามารถในการทำซ้ำ (การกระทำด้วยตนเอง) หน้าที่หลักของ DNA คือการจัดเก็บข้อมูลและการถ่ายโอนข้อมูลทางพันธุกรรมซึ่งบันทึกไว้ในรูปแบบของลำดับนิวคลีโอไทด์ ความเสถียรของโมเลกุล DNA นั้นได้รับการบำรุงรักษาเนื่องจากระบบการชดเชยที่มีประสิทธิภาพ (การกู้คืน) แต่ถึงแม้ว่าพวกเขาจะไม่สามารถกำจัดผลกระทบที่ไม่พึงประสงค์ที่นำไปสู่การกลายพันธุ์ในท้ายที่สุด DNA Eukaryotic Cells มีความเข้มข้นในเคอร์เนล Mitochondria และ Plastids และ Prokaryotic - ตั้งอยู่โดยตรงในไซโตพลาสซึม Nuclear DNA เป็นพื้นฐานของโครโมโซมมันเป็นตัวแทนของโมเลกุลที่ไม่ได้แยกออก DNA mitochondria, Plastid และ Prokaryotic มีรูปร่างเป็นวงแหวน
กรด ribonucleic (RNA) - Biopolymer ซึ่งโมโนเมอร์เป็น ribonucleotides พวกเขายังมีฐานไนโตรเจนสี่ตัว - Adenine (A), Uracil (Y), Guanine (G) หรือ Cytosine (C) ซึ่งแตกต่างจาก DNA บนหนึ่งในพื้นที่ (แทนที่จะเป็น Tymina ใน RNA ใน RNA, uracil meet) สารตกค้างของการเพนกวากน้ำตาลใน ribonucleotides เป็นตัวแทนของ Ribose RNA เป็นโมเลกุลเดี่ยวที่ควั่นด้วยข้อยกเว้นของไวรัสบางอย่าง สามประเภทหลักของ RNA ถูกแยกได้: ข้อมูลหรือเมทริกซ์ (IRNA, MRNA), Ribosomal (RRNA) และการขนส่ง (TRNA) พวกเขาทั้งหมดเกิดขึ้นในกระบวนการ การถอดความ - เขียนใหม่ด้วยโมเลกุล DNA
และRNA เป็นเศษส่วน RNA ที่เล็กที่สุดในเซลล์ (2-4%) ซึ่งได้รับการชดเชยจากความหลากหลายของพวกเขาเนื่องจาก IRNN ที่แตกต่างกันหลายพันตัวอาจมีอยู่ในเซลล์เดียว เหล่านี้เป็นโมเลกุลเดี่ยวที่กีดขวางซึ่งเป็นเมทริกซ์สำหรับการสังเคราะห์โซ่โพลีเปปไทด์ ข้อมูลเกี่ยวกับโครงสร้างของโปรตีนถูกบันทึกไว้ในรูปแบบของลำดับนิวคลีโอไทด์และนิวคลีโอไทด์แฝดจะถูกเข้ารหัสแต่ละกรดอะมิโน - codon.
rRNA เป็น RNA จำนวนมากที่สุดในเซลล์ (สูงถึง 80%) น้ำหนักโมเลกุลของพวกเขาเฉลี่ย 3,000-5,000; พวกเขาถูกสร้างขึ้นในนิวเคลียสและรวมอยู่ในช่องว่างของเซลล์ - ไรโบโซม RRNA เห็นได้ชัดว่ามีบทบาทในกระบวนการสังเคราะห์โปรตีน
ต.RNA เป็นโมเลกุลที่เล็กที่สุดของ RNA เนื่องจากมีเพียง 73-85 นิวคลีโอไทด์ สัดส่วนของจำนวนเซลล์ RNA ทั้งหมดประมาณ 16% ฟังก์ชั่น TRNA - การขนส่งกรดอะมิโนไปยังสถานที่สังเคราะห์โปรตีน (บน Ribosomes) ในรูปแบบของโมเลกุล Trna มีลักษณะคล้ายใบไม้โคลเวอร์ ที่ปลายด้านหนึ่งโมเลกุลเป็นพล็อตสำหรับการติดกรดอะมิโนและในหนึ่งในลูป - นิวคลีโอไทด์แฝด, Codon เสริมของ IRNK และการพิจารณาซึ่งกรดอะมิโนจะถูกโอนไปยัง TRNA - antikodon

RNA ทุกประเภทใช้ส่วนที่ใช้งานอยู่ในกระบวนการดำเนินการทางพันธุกรรมซึ่งถูกเขียนใหม่จาก DNA ไปยัง IRNA และการสังเคราะห์โปรตีนจะดำเนินการในภายหลัง TRNA ในกระบวนการสังเคราะห์โปรตีนให้กรดอะมิโนเป็นไรโบโซมและ RRNA เป็นส่วนหนึ่งของ Ribosomes โดยตรง
Adenosine Trifosphoric Acid (ATP) - นี่คือนิวคลีโอไทด์ที่มีอยู่นอกเหนือไปจากฐานของ adenine และสารตกค้างของ ribosa, สามตกค้างของกรดฟอสฟอริก การเชื่อมโยงระหว่างด๊ดฟอสเฟตที่มีต่อฟอสเฟตเมื่อเร็ว ๆ นี้คือ macroeergic (ด้วยความแตกแยก, 42 kJ / mol ของพลังงานมีความแตกต่าง) ในขณะที่พันธะเคมีมาตรฐานที่มีการแยกให้ 12 kJ / mol หากจำเป็นการสื่อสารแบบ macroeergic ของ ATP คือการแยกกรด adenosine infusion (ADP) สารตกค้างฟอสฟอริกและพลังงานมีความแตกต่าง:
ATP + H 2 O $ → $ ADF + H 3 PO 4 + 42 KJ
ADP ยังสามารถฉลาดได้ด้วยการก่อตัวของแอมป์ (กรด monophosphoric adenosine) และสารตกค้างของกรดฟอสฟอริก:
ADF + H 2 O $ → $ AMP + H 3 PO 4 + 42 KJ
ในกระบวนการของการแลกเปลี่ยนพลังงาน (ในการหายใจการหมัก) เช่นเดียวกับในกระบวนการของการสังเคราะห์ด้วยแสง ADF ยึดติดกับสารตกค้างฟอสฟอรัสและกลายเป็น ATP ปฏิกิริยาการกู้คืน ATP เรียกว่า phosphorylation. ATP เป็นแหล่งพลังงานสากลสำหรับกระบวนการทั้งหมดของกิจกรรมสำคัญของสิ่งมีชีวิต
การศึกษาองค์ประกอบทางเคมีของเซลล์ของสิ่งมีชีวิตทั้งหมดแสดงให้เห็นว่าพวกเขามีองค์ประกอบทางเคมีเดียวกัน สารเคมีดำเนินการฟังก์ชั่นเดียวกัน นอกจากนี้ส่วน DNA ที่ย้ายจากร่างกายหนึ่งไปยังอีกส่วนหนึ่งจะทำงานในนั้นและโปรตีนสังเคราะห์จากแบคทีเรียหรือเห็ดจะทำหน้าที่ของฮอร์โมนหรือเอนไซม์ในร่างกายมนุษย์ นี่เป็นหนึ่งในหลักฐานของความสามัคคีของต้นกำเนิดของโลกอินทรีย์

โครงสร้างเซลล์ ความสัมพันธ์ของโครงสร้างและหน้าที่ของชิ้นส่วนและเซลล์อิงเกณฑ์ - พื้นฐานของความซื่อสัตย์
โครงสร้างเซลล์
โครงสร้างของเซลล์ Prokaryotic และ Eukaryotic
ส่วนประกอบโครงสร้างหลักของเซลล์คือเมมเบรนพลาสม่าไซโตพลาสซึมและอุปกรณ์ทางพันธุกรรม ขึ้นอยู่กับลักษณะขององค์กรเซลล์ชนิดหลักสองประเภทมีความโดดเด่น: Prokaryotic และ Eukaryotic ความแตกต่างที่สำคัญของเซลล์ prokaryotic จาก Eukaryotic คือองค์กรของเครื่องมือทางพันธุกรรมของพวกเขา: มันอยู่ในไซโตพลาสซึมโดยตรง (พื้นที่ของไซโตพลาสซึมนี้เรียกว่า นิวเคลียส) และไม่แยกออกจากโครงสร้างของเมมเบรนในขณะที่ Eukaryotes เป็นส่วนที่โหดร้ายของ DNA ที่เข้มข้นในแกนกลางล้อมรอบด้วยเมมเบรนคู่ นอกจากนี้ข้อมูลทางพันธุกรรมของเซลล์ prokaryotic ในนิวเคลียสบันทึกในโมเลกุล DNA ของวงแหวนและโมเลกุล DNA ถูกปลดล็อคใน Eukaryot
ในทางตรงกันข้ามกับ Eukaryotes ไซโตพลาสซึมของเซลล์ prokaryotic ยังมีปริมาณอวัยวะขนาดเล็กในขณะที่ Eukaryotic ซึ่งเป็นสิ่งที่มีความหลากหลายของโครงสร้างเหล่านี้มีลักษณะ
โครงสร้างและหน้าที่ของเยื่อหุ้มชีวภาพ
โครงสร้าง Biomembrane เยื่อหุ้มเซลล์ จำกัด เซลล์และช่องเสียบเมมเบรนของเซลล์ยูคาริโอตมีองค์ประกอบทางเคมีและโครงสร้างทั้งหมด พวกเขามีไขมันโปรตีนและคาร์โบไฮเดรต ไขมันของเมมเบรนจะแสดงส่วนใหญ่โดยฟอสโฟไลปิดและคอเลสเตอรอล โปรตีน Membranes ส่วนใหญ่เป็นของโปรตีนที่ซับซ้อนเช่น Glycoproteins คาร์โบไฮเดรตไม่พบในเมมเบรนด้วยตัวเองพวกเขาเกี่ยวข้องกับโปรตีนและไขมัน ความหนาของเมมเบรนคือ 7-10 นาโนเมตร

ตามรูปแบบโมเสคของเหลวที่ยอมรับในปัจจุบันของโครงสร้างของเมมเบรนไขมันในรูปแบบสองชั้นหรือ lipid bisoyeซึ่งซึ่ง "หัว" ของ Hydrophilic ของโมเลกุลไขมันจะถูกเปิดออกไปด้านนอกและ "หาง" ที่ไม่ชอบน้ำถูกซ่อนไว้โดยเมมเบรน "หาง" เหล่านี้เนื่องจากความสะอาดของมันให้การแยกเฟสของสภาพอากาศภายในเซลล์และสภาพแวดล้อมของมัน ไขมันที่มีการโต้ตอบหลายประเภทคือโปรตีน ส่วนหนึ่งของโปรตีนตั้งอยู่บนพื้นผิวของเมมเบรน โปรตีนดังกล่าวเรียกว่า อุปกรณ์ต่อพ่วง, หรือ พื้นผิว. โปรตีนอื่น ๆ บางส่วนหรือแช่อยู่ในเมมเบรน - นี้ เป็นส่วนหนึ่ง หรือ โปรตีนที่แช่. โปรตีนเมมเบรนดำเนินการโครงสร้างการขนส่งตัวเร่งปฏิกิริยาตัวรับและฟังก์ชั่นอื่น ๆ
เมมเบรนไม่เหมือนกับคริสตัลส่วนประกอบของพวกเขากำลังเคลื่อนไหวอยู่ตลอดเวลาเนื่องจากการหยุดพักระหว่างโมเลกุลไขมันซึ่งสารต่าง ๆ สามารถตกอยู่ในเซลล์
เยื่อบุชีวภาพแตกต่างกันไปตามตำแหน่งในเซลล์องค์ประกอบทางเคมีและฟังก์ชั่นที่ดำเนินการ เมมเบรนประเภทหลักคือพลาสมาและภายใน เมมเบรนพลาสม่า มีไขมันประมาณ 45% (รวมถึง glycolipids), โปรตีน 50% และคาร์โบไฮเดรต 5% โซ่คาร์โบไฮเดรตที่เป็นส่วนหนึ่งของโปรตีน - glycoproteins ที่ซับซ้อนและ lipids-glycolipids ที่ซับซ้อนยื่นออกมาเหนือพื้นผิวของเมมเบรน Plasmalemma Glycoproteins มีความเฉพาะเจาะจงมาก ตัวอย่างเช่นมีการรับรู้ซึ่งกันและกันของเซลล์ที่มีอยู่รวมถึงสปิมโตะและไข่
บนพื้นผิวของเซลล์สัตว์โซ่คาร์โบไฮเดรตสร้างชั้นพื้นผิวบาง ๆ - glycocalix มันถูกเปิดเผยในเซลล์สัตว์เกือบทั้งหมด แต่ระดับความรุนแรงของ Non-Etinakov (10-50 ไมครอน) Glycocalix ให้การเชื่อมต่อเซลล์โดยตรงกับสภาพแวดล้อมภายนอกมีการย่อยอาหารนอกเซลล์ Glycicalce มีตัวรับ เซลล์ของแบคทีเรียพืชและเห็ดนอกเหนือไปจากพลาสมุนามาล้อมรอบด้วยกระสุนเซลล์
เมมเบรนภายใน เซลล์ Eukaryotic คั่นส่วนต่าง ๆ ของเซลล์การสร้าง "ช่อง" ที่แปลกประหลาด - ผู้ให้ความสามารถซึ่งก่อให้เกิดการแยกกระบวนการเผาผลาญและพลังงานต่างๆ พวกเขาอาจแตกต่างกันไปตามองค์ประกอบทางเคมีและฟังก์ชั่นที่ดำเนินการ แต่โครงสร้างทั่วไปของโครงสร้างจะถูกเก็บรักษาไว้
ฟังก์ชั่นเมมเบรน:
- จำกัด มันอยู่ในความจริงที่ว่าพวกเขาแยกพื้นที่ภายในของเซลล์จากสภาพแวดล้อมภายนอก เมมเบรนสามารถซึมผ่านได้อย่างอิสระเท่านั้นที่จะเอาชนะเฉพาะสารเหล่านั้นที่เซลล์ต้องการในขณะที่มีกลไกสำหรับการขนส่งสารสำคัญ
- ตัวรับ มันถูกผูกไว้กับการรับรู้สัญญาณสิ่งแวดล้อมและการส่งข้อมูลนี้ภายในเซลล์ โปรตีนตัวรับพิเศษมีหน้าที่รับผิดชอบในฟังก์ชั่นนี้ โปรตีนเมมเบรนมีหน้าที่รับผิดชอบต่อการรับรู้ของมือถือตามหลักการของ "คนแปลกหน้าของพวกเขาเอง" รวมถึงการก่อตัวของสารประกอบระหว่างเซลล์การศึกษามากที่สุดที่มีการซินเป็นเซลล์ประสาท
- ตัวเร่งปฏิกิริยา เมมเบรนเป็นคอมเพล็กซ์เอนไซม์จำนวนมากเนื่องจากกระบวนการสังเคราะห์ที่มีความเข้มข้นเกิดขึ้น
- เปลี่ยนพลังงาน มันเกี่ยวข้องกับการก่อตัวของพลังงานความเข้มของมันในรูปแบบของ ATP และการใช้จ่าย
- การเสริม เมมเบรนนั้นแตกต่างจากพื้นที่ภายในเซลล์จึงแยกสารเริ่มต้นของปฏิกิริยาและเอนไซม์ที่สามารถทำปฏิกิริยาได้
- การก่อตัวของผู้ติดต่อระหว่างเซลล์ แม้จะมีความจริงที่ว่าความหนาของเมมเบรนมีขนาดเล็กมากจนเป็นไปไม่ได้ที่จะแยกแยะด้วยตาเปล่ามันในมือข้างหนึ่งมีอุปสรรคที่ค่อนข้างน่าเชื่อถือสำหรับไอออนและโมเลกุลโดยเฉพาะอย่างยิ่งน้ำที่ละลายน้ำได้และอื่น ๆ มันให้การถ่ายโอนของพวกเขาไปยังเซลล์และออกไปด้านนอก
- ขนส่ง.

การขนส่งเมมเบรน เนื่องจากความจริงที่ว่าเซลล์เป็นระบบชีวภาพระดับประถมศึกษาจึงเปิดระบบเพื่อให้แน่ใจว่าการเผาผลาญและพลังงานการบำรุงรักษาสภาวะสมดุลการเจริญเติบโตหงุดหงิดและกระบวนการอื่น ๆ ต้องมีการถ่ายโอนสารผ่านเมมเบรน - การขนส่งเมมเบรน ปัจจุบันการขนส่งสารผ่านเมมเบรนเซลล์จะถูกแบ่งออกเป็นแอคทีฟเรื่อย ๆ endo- และ exocytosis
การขนส่งแบบพาสซีฟ - นี่คือประเภทของการขนส่งที่เกิดขึ้นโดยไม่มีค่าใช้จ่ายพลังงานจากความเข้มข้นที่มากขึ้นไปจนถึงขนาดเล็ก โมเลกุลที่ไม่ใช่ขั้วขนาดเล็กที่ละลายในไขมันในไขมัน (O 2, CO 2) แทรกซึมเข้าไปในกรงได้อย่างง่ายดาย การแพร่กระจายอย่างง่าย. ไม่ละลายในไขมันรวมถึงอนุภาคขนาดเล็กที่มีประจุจะถูกหยิบขึ้นมาโดย perecenisters โปรตีนหรือผ่านช่องพิเศษ (กลูโคส, กรดอะมิโน, K +, PO 4 3-) การขนส่งแบบพาสซีฟประเภทนี้เรียกว่า การแพร่กระจายแสง. น้ำเข้าสู่กรงผ่านรูขุมขนในเฟสไขมันรวมถึงช่องพิเศษที่เรียงรายไปด้วยโปรตีน การขนส่งทางน้ำผ่านเมมเบรนเรียกว่า osmosis

Osmosis เป็นสิ่งสำคัญอย่างยิ่งในชีวิตของเซลล์ตั้งแต่หากมีการวางโซลูชันที่มีความเข้มข้นของน้ำเค็มสูงกว่าในการแก้ปัญหาเซลล์น้ำจะเริ่มออกจากเซลล์และปริมาณการใช้ชีวิตจะเริ่มลดลง ในเซลล์สัตว์เซลล์ถูกกวาดโดยทั่วไปและในผัก - ความล่าช้าหลังไซโตพลาสซึมจากผนังเซลล์ซึ่งเรียกว่า plasmolysis. เมื่อวางเซลล์ให้เข้มข้นน้อยกว่าพลาสซึมน้อยกว่าการแก้ปัญหาการขนส่งทางน้ำเกิดขึ้นในทิศทางตรงกันข้ามกับเซลล์ อย่างไรก็ตามมีขีด จำกัด แรงดึงของเมมเบรนไซโตพลาสซึมและเซลล์สัตว์ในที่สุดก็ระเบิดและโรงงานไม่อนุญาตให้ใช้ผนังเซลล์ที่ทนทานในโรงงาน ปรากฏการณ์ของการเติมเนื้อหามือถือของพื้นที่ภายในทั้งหมดของเซลล์เรียกว่า การทำให้งงงวย. ความเข้มข้นของเซลล์ของเกลือควรพิจารณาในการเตรียมยาโดยเฉพาะอย่างยิ่งสำหรับการบริหารทางหลอดเลือดดำเนื่องจากสามารถนำไปสู่ความเสียหายต่อเซลล์เม็ดเลือด (สำหรับสิ่งนี้น้ำเกลือจะใช้กับความเข้มข้นของโซเดียมคลอไรด์ 0.9%) สิ่งนี้มีความสำคัญเท่าเทียมกันในการเพาะปลูกของเซลล์และเนื้อเยื่อรวมถึงอวัยวะสัตว์และพืช

การขนส่งที่ใช้งานอยู่ มันเกิดขึ้นกับค่าใช้จ่ายของพลังงาน ATP จากความเข้มข้นที่ต่ำกว่าของสารมากขึ้น มันดำเนินการด้วยความช่วยเหลือของโปรตีนปั๊มพิเศษ โปรตีนจะถูกสูบผ่านเมมเบรนไอออน K +, NA +, CA 2+ และอื่น ๆ ซึ่งก่อให้เกิดการขนส่งสารอินทรีย์ที่จำเป็นเช่นเดียวกับการเกิดแรงกระตุ้นของเส้นประสาท ฯลฯ
endocytosis - นี่เป็นกระบวนการดูดซับสารที่ใช้งานอยู่โดยเซลล์ซึ่งเป็นเมมเบรนรูปแบบเงินบำนาญจากนั้นสร้างฟองเมมเบรน - fagosomesซึ่งสรุปวัตถุที่ดูดซับได้ จากนั้น lysosome หลักจะถูกรวมเข้ากับ phageosoma และรูปแบบ lysosoma รอง, หรือ phagolizosoma, หรือ เครื่องถ่านย่อยอาหาร. เนื้อหาของฟองสบู่ cleaves เอนไซม์ของ lysosomes และผลิตภัณฑ์ที่แตกแยกถูกดูดซับและดูดซับโดยเซลล์ สารตกค้างที่ไม่ทาสีจะถูกลบออกจากเซลล์โดย exocytosis endocytosis มีสองประเภทหลัก: phagocytosis และ pinocytosis
phagocytosis - นี่คือกระบวนการของการจับภาพพื้นผิวเซลล์และการดูดซับโดยเซลล์ของอนุภาคของแข็งและ pinocytosis - ของเหลว Phagocytosis ดำเนินการส่วนใหญ่ในเซลล์สัตว์ (สัตว์เซลล์เดียว, เม็ดเลือดขาวของมนุษย์) มันให้สารอาหารของพวกเขาและบ่อยครั้งที่การปกป้องร่างกาย Pinocytosis ถูกดูดซับโดยโปรตีน, แอนติเจน - แอนติบอดีคอมเพล็กซ์ในกระบวนการของปฏิกิริยาภูมิคุ้มกัน ฯลฯ อย่างไรก็ตามไวรัสจำนวนมากยังตกอยู่ในเซลล์โดย pinocytosis หรือ phagocytosis ในเซลล์ของพืชและเห็ด Phagocytosis แทบจะเป็นไปไม่ได้เพราะล้อมรอบด้วยเปลือกเซลล์ที่แข็งแรงทนทาน

การเสียดสี - กระบวนการ endocytosis ผกผัน ดังนั้นอาหารที่ตกค้างที่ไม่ได้รับการด๊ดจากฉุกเฉินทางเดินอาหารจึงมีความโดดเด่นเซลล์และสิ่งมีชีวิตโดยรวมเป็นสิ่งจำเป็นสำหรับกิจกรรมที่สำคัญ ตัวอย่างเช่นการถ่ายโอนพัลส์เส้นประสาทเกิดขึ้นเนื่องจากการเลือกตัวกลางทางเคมีโดยการปล่อยชีพจร - ผู้ไกล่เกลี่ยและในเซลล์พืชคาร์โบไฮเดรตเสริมของเปลือกเซลล์มีความโดดเด่น
เปลือกเซลล์ของเซลล์พืชเห็ดและแบคทีเรีย ด้านนอกเมมเบรนเซลล์สามารถเน้นเฟรมที่ทนทาน - เปลือกเซลล์ หรือ ผนังเซลล์.
ในพืชฐานของเปลือกเซลล์คือ เซลลูโลสบรรจุในการรวมกลุ่มของโมเลกุล 50-100 ช่องว่างระหว่างพวกเขาเต็มไปด้วยน้ำและคาร์โบไฮเดรตอื่น ๆ เปลือกของกรงพืชถูกแทรกซึมโดย tubules - plasmodesmaซึ่งมีเมมเบรนของเครือข่าย endoplasmic กำลังผ่านไป ตาม Plasmodems สารขนส่งจะถูกดำเนินการระหว่างเซลล์ อย่างไรก็ตามสารขนส่งเช่นน้ำสามารถเกิดขึ้นได้บนผนังเซลล์เอง เมื่อเวลาผ่านไปสารต่าง ๆ จะสะสมในพืชในพืชรวมถึง Tubyl หรือคล้ายน้ำมันซึ่งนำไปสู่การไวต่อการไวด์หรือการอำนวยความสะดวกของผนังเซลล์เองแทนที่น้ำและการกำจัดเนื้อหามือถือ ระหว่างผนังเซลล์ของเซลล์ที่อยู่ใกล้เคียงจะอยู่ปะเก็นคล้ายวุ้น - แผ่นมัธยฐานที่ยึดไว้ระหว่างตัวเองและซีเมนต์ร่างกายของพืชโดยรวม พวกเขาถูกทำลายเฉพาะในกระบวนการของการสุกของผลไม้และเมื่อตกลงมาจากใบไม้

ผนังเซลล์ของเซลล์เห็ดจะเกิดขึ้น ไคติน - คาร์โบไฮเดรตที่มีไนโตรเจน พวกเขาแข็งแรงพอและเป็นโครงกระดูกเซลล์ภายนอก แต่ยังคงอยู่ในพืชป้องกัน phagocytosis
แบคทีเรียในผนังเซลล์มีคาร์โบไฮเดรตที่มีชิ้นส่วนเปปไทด์ - mareinอย่างไรก็ตามเนื้อหานั้นแตกต่างกันอย่างมีนัยสำคัญในกลุ่มแบคทีเรียที่แตกต่างกัน โพลีแซคคาไรด์อื่น ๆ สร้างแคปซูลเมือกการปกป้องแบคทีเรียจากอิทธิพลภายนอกอาจถูกจัดสรรที่ด้านบนของผนังเซลล์
เปลือกเป็นตัวกำหนดรูปร่างของเซลล์ทำหน้าที่เป็นการสนับสนุนเชิงกลให้ทำงานฟังก์ชั่นป้องกันให้คุณสมบัติออสโมติกของเซลล์ จำกัด การยืดปริมาณการใช้ชีวิตและป้องกันการเพิ่มแรงดึงของเซลล์แรงดึงเนื่องจากการไหลของน้ำ นอกจากนี้ผนังเซลล์จะเอาชนะน้ำและละลายในสารมันก่อนเข้าสู่ไซโตพลาสซึมหรือในทางตรงกันข้ามเมื่อออกจากมันในขณะที่น้ำถูกส่งเร็วกว่าพลาสซึม
พลาสซึม
พลาสซึม - นี่คือเนื้อหาภายในของเซลล์ Cellides ทั้งหมดของเซลล์หลักและผลิตภัณฑ์ที่สำคัญต่าง ๆ จะถูกส่งเข้ามา
ไซโตพลาสซึมเชื่อมโยงทุกส่วนของเซลล์ในหมู่ตัวเองปฏิกิริยาการเผาผลาญจำนวนมากไหลอยู่ในนั้น ไซโตพลาสซึมถูกแยกออกจากสภาพแวดล้อมและแบ่งออกเป็นฝูงเมมเบรนนั่นคือเซลล์มีอยู่ในโครงสร้างเมมเบรน มันสามารถอยู่ในสองรัฐ - Zola และเจล โซล - นี่คือสถานะกึ่งของเหลวออกซิเจนของไซโตพลาสซึมซึ่งกระบวนการของกิจกรรมสำคัญดำเนินการอย่างเข้มข้นมากที่สุดและ เจล - รัฐที่มีลักษณะที่หนาแน่นมากขึ้นที่ขัดขวางการไหลของปฏิกิริยาเคมีและสารขนส่ง
ส่วนที่เป็นของเหลวของไซโตพลาสซึมที่ไม่มีออแกไนซ์เรียกว่า hyaloplasm. Hyaloplasma หรือ Cytosol เป็นโซลูชันคอลลอยด์ที่ระบบกันสะเทือนที่แปลกประหลาดมีอนุภาคขนาดใหญ่เพียงพอเช่นโปรตีนที่ล้อมรอบด้วยโมเลกุลของน้ำ การสะสมของการระงับนี้ไม่เกิดขึ้นเนื่องจากความจริงที่ว่าพวกเขามีประจุเดียวกันและขับไล่จากกันและกัน
organoid
organoid - เหล่านี้เป็นส่วนประกอบของเซลล์ถาวรที่ทำหน้าที่บางอย่าง
ขึ้นอยู่กับลักษณะของโครงสร้างพวกเขาแบ่งออกเป็นเมมเบรนและไม่ใช่ตราสัญลักษณ์ เมมเบรน Organoisians ในทางกลับกันอ้างถึงเดี่ยวหรี่แสงได้ (เครือข่าย endoplasmic, Golgi และ Lizosome Complex) หรือสองขูด (mitochondria, plastids และเคอร์เนล) ไม่ทุบ Organoisses เป็น Ribosomes, Microtubule, Microfilaments และ Cellular Center Prokaryotam จาก Organoids ที่ระบุไว้มีอยู่ในไรโบโซมเท่านั้น
โครงสร้างและหน้าที่ของเคอร์เนล แกนกลาง - อริกอริกอยขนาดใหญ่ขูดสองขูดนอนอยู่ตรงกลางของเซลล์หรือบนรอบนอก ขนาดของเคอร์เนลสามารถแตกต่างกันภายใน 3-35 ไมครอน รูปร่างของนิวเคลียสมักจะเป็นทรงกลมหรือวงรี แต่ยังมีรูปทรงกลม, แกนหมุน, รูปร่าง beobovo, ใบมีดและแม้กระทั่งเมล็ดที่แบ่งส่วน นักวิจัยบางคนเชื่อว่ารูปร่างของนิวเคลียสสอดคล้องกับรูปแบบของเซลล์เอง
เซลล์ส่วนใหญ่มีหนึ่งหลัก แต่อาจมีเซลล์ตับและหัวใจสองเซลล์และในจำนวนของเซลล์ประสาท - มากถึง 15 เส้นใยของกล้ามเนื้อโครงร่างมักจะมีนิวเคลียสจำนวนมาก แต่พวกเขาไม่ใช่เซลล์ในแบบเต็ม ความรู้สึกของคำว่าเนื่องจากมีการเกิดขึ้นจากการควบรวมกิจการของเซลล์หลายเซลล์
เคอร์เนลล้อมรอบไป เปลือกนิวเคลียร์และอวกาศภายในของมันเต็มไปด้วย น้ำนิวเคลียร์, หรือ nucleoplasm (Karyoplasm)ที่คุณจัดส่ง chromatin และ nadryshko. เคอร์เนลแสดงฟังก์ชั่นที่จำเป็นเช่นการจัดเก็บและถ่ายโอนข้อมูลทางพันธุกรรมรวมถึงการควบคุมเซลล์ที่สำคัญของเซลล์

บทบาทของเคอร์เนลในการถ่ายโอนข้อมูลทางพันธุกรรมได้รับการพิสูจน์อย่างมั่นใจในการทดลองกับสาหร่าย acetabulary สีเขียว ในเซลล์ยักษ์เดียวที่มีความยาว 5 ซม. แยกหมวกขาและ rhizoid ในเวลาเดียวกันมันมีเพียงเคอร์เนลเดียวที่ตั้งอยู่ใน Rhizoid ในช่วงทศวรรษที่ 1930 I. Hemmerling ปลูกเมล็ดเคอร์เนลของ acetabulary ชนิดหนึ่งที่มีสีเขียวใน Rhizoid ของสปีชีส์อีกชนิดหนึ่งมีสีน้ำตาลจากที่เคอร์เนลถูกลบ หลังจากผ่านไประยะหนึ่งหมวกใหม่ที่ปลูกที่โรงงานด้วยแกนปลูกถ่ายเหมือนสาหร่ายเคอร์เนล ในเวลาเดียวกันหมวกหรือนิวเคลียสที่มีอยู่ในขาแยกออกจาก Rhizoid ไม่ได้อยู่หลังจากเวลาที่พวกเขาเสียชีวิต

เปลือกนิวเคลียร์ เกิดจากสองเยื่อหุ้มสอง - ด้านนอกและด้านในระหว่างที่มีพื้นที่ว่าง พื้นที่ intermogram ถูกรายงานไปยังโพรงของเครือข่าย endoplasmic หยาบและเมมเบรนด้านนอกของนิวเคลียสสามารถพกของไรโบโซม ปลอกนิวเคลียร์ถูกแทรกซึมด้วยรูขุมขนจำนวนมากขอบมีโปรตีนพิเศษ ผ่านรูขุมขนมีการขนส่งสาร: โปรตีนที่จำเป็น (รวมถึงเอนไซม์), ไอออน, นิวคลีโอไทด์และสารอื่น ๆ ตกอยู่ในเคอร์เนลและโมเลกุลของ RNA ที่ใช้งานโปรตีนย่อยของไรโบโซม ดังนั้นฟังก์ชั่นของเปลือกนิวเคลียร์คือเนื้อหาของเคอร์เนลของไซโตพลาสซึมเช่นเดียวกับการควบคุมการเผาผลาญระหว่างแกนกลางและไซโตพลาสซึม
นิวเคลียสเรียกเนื้อหาของเคอร์เนลที่ Chromatin และนิวเคลียสถูกจัดส่ง มันเป็นสารละลายคอลลอยด์โดยองค์ประกอบทางเคมีที่มีลักษณะคล้ายพลาสซึม เอนไซม์นิวเคลียสตัวเร่งปฏิกิริยาการแบ่งปันกรดอะมิโนนิวคลีโอไทด์โปรตีนและนิวคลีโอพลาสซึมอื่น ๆ มีความเกี่ยวข้องกับ Hyaloplasm ผ่านรูขุมขนนิวเคลียร์ ฟังก์ชั่น Nucleoplasm เช่นเดียวกับ Hyaloplasms คือเพื่อให้แน่ใจว่าความสัมพันธ์ขององค์ประกอบโครงสร้างทั้งหมดของเคอร์เนลและการใช้งานของปฏิกิริยาของเอนไซม์จำนวนมาก
chromatin เรียกว่าการรวมกันของเธรดบางและเม็ดที่แช่อยู่ในนิวเคลียส เป็นไปได้ที่จะระบุมันเฉพาะเมื่อทาสีเนื่องจากดัชนีการหักเหของ Chromatin และ Nucleoplasm อยู่ที่ประมาณเดียวกัน ส่วนประกอบของ Chromatin Nitched เรียกว่า eukhromatinและเม็ด - heterochromatin. Euchromatin ถูกปิดผนึกไม่ดีเนื่องจากข้อมูลทางพันธุกรรมถูกอ่านจากในขณะที่ heterochromatin ที่หมุนวนมากขึ้นนั้นไม่ได้ใช้งานทางพันธุกรรม
Chromatin เป็นการปรับโครงสร้างของโครโมโซมในเคอร์เนลที่มีรอยบาก ดังนั้นโครโมโซมจึงมีอยู่ในเคอร์เนลเพียงอย่างต่อเนื่องเพียงการเปลี่ยนแปลงของรัฐขึ้นอยู่กับฟังก์ชั่นที่เคอร์เนลทำงานในขณะนี้
Chromatin ส่วนใหญ่รวมถึงโปรตีน-nucleoproteins (deoxyribonucleoproteins และ ribonucleoproteins) เช่นเดียวกับเอนไซม์ที่สำคัญที่สุดที่เกี่ยวข้องกับการสังเคราะห์การสังเคราะห์กรดนิวเคลียสและสารอื่น ๆ บางชนิด
ฟังก์ชั่นของ Chromatin ประกอบด้วยประการแรกในการสังเคราะห์กรดนิวคลีอิกที่เฉพาะเจาะจงเฉพาะสำหรับสิ่งมีชีวิตนี้ซึ่งนำการสังเคราะห์โปรตีนเฉพาะและประการที่สองในการถ่ายโอนคุณสมบัติทางพันธุกรรมจากเซลล์แม่ซึ่งเป็นเส้นด้ายโครมาตินที่บรรจุในโครโมโซม .
nadryshko - ทรงกลมมองเห็นได้อย่างดีภายใต้กล้องจุลทรรศน์ Caller ที่มีขนาดเส้นผ่าศูนย์กลาง 1-3 μm มันเกิดขึ้นในส่วนของ Chromatin ซึ่งข้อมูลเกี่ยวกับโครงสร้างของ RDNA และ Ribosomes ถูกเข้ารหัส นิวเคลียสในแกนกลางมักเป็นหนึ่งในเซลล์ที่มีกระบวนการอย่างเข้มข้นของกิจกรรมที่สำคัญเกิดขึ้นนิวเคลียสอาจเป็นสองหรือมากกว่านั้น ฟังก์ชั่นของนิวเคลียส - การสังเคราะห์ RRNA และการประกอบของ Subunits Ribosomes โดยการรวม RRNA กับโปรตีนที่มาจากไซโตพลาสซึม
mitochondria - ออร์แกอร์อิดสองหวีของรูปทรงกลมวงรีหรือกลิ้งแม้ว่าพวกเขาจะตอบสนองและเกลียว (ในอสุจิ) เส้นผ่านศูนย์กลางของไมโตคอนเดรียสูงถึง 1 μmและความยาวสูงถึง 7 ไมครอน พื้นที่ภายใน Mitochondria เต็มไปด้วยเมทริกซ์ เมทริกซ์ - นี่คือสารหลักของไมโตคอนเดรีย โมเลกุล DNA แบบวงแหวนและไรโบสโมเลกุลจะถูกแช่อยู่ในนั้น Mitochondria เมมเบรนกลางแจ้งเรียบมันไม่สามารถเข้าใจได้สำหรับสารหลายชนิด เมมเบรนภายในได้เติบโตขึ้น - ศึกเพิ่มพื้นที่ผิวของเมมเบรนเพื่อการไหลของปฏิกิริยาเคมี คอมเพล็กซ์โปรตีนจำนวนมากตั้งอยู่บนพื้นผิวของเมมเบรนซึ่งเป็นห่วงโซ่ระบบทางเดินหายใจเช่นเดียวกับเอนไซม์ ATP-Synthetase เห็ด ใน Mitochondria เวทีแอโรบิกของการหายใจไหลในระหว่างการสังเคราะห์ ATP เกิดขึ้น

ลูกแก้ว - มีลักษณะเฉพาะตัวสองปูขนาดใหญ่สำหรับเซลล์ผักเท่านั้น Space Internal Plastid ที่เต็มไปด้วย stroma, หรือ เมทริกซ์. ใน stroma มีระบบฟองเมมเบรนที่พัฒนาขึ้นหรือน้อยกว่า - tylakoidsใครถูกรวบรวมในสแต็ค - graarsเช่นเดียวกับโมเลกุล DNA ของตัวเองและ Ribosomes มีพลาสติกหลักสี่ประเภท: Chloroplasts, Chromoplasts, Leukoplasts และตกตะกอน
คลอโรพลาสต์ - เหล่านี้เป็นพลาสต์สีเขียวที่มีขนาดเส้นผ่าศูนย์กลาง 3-10 μmแยกได้อย่างดีภายใต้กล้องจุลทรรศน์ พวกเขามีเฉพาะในส่วนสีเขียวของพืช - ใบลำต้นอ่อนดอกไม้และผลไม้ Chloroplasts ส่วนใหญ่มีรูปไข่หรือรูปไข่แบบวงรี แต่อาจมีการเติมพลังเกลียวและใบมีด ปริมาณของคลอโรพลาสต์ในเซลล์โดยเฉลี่ยอยู่ในช่วง 10 ถึง 100 ชิ้น อย่างไรก็ตามตัวอย่างเช่นสาหร่ายบางคนสามารถอยู่คนเดียวมีขนาดที่สำคัญและรูปร่างที่ซับซ้อน - แล้วก็เรียกว่า chromatorm. ในกรณีอื่น ๆ จำนวนคลอโรพลาสต์สามารถเข้าถึงได้หลายร้อยในขณะที่ขนาดของพวกเขามีขนาดเล็ก ระบายสีคลอโรพลาสต์เนื่องจากการสังเคราะห์ด้วยแสงเม็ดสีหลัก - คลอโรฟิลล์แม้ว่าพวกเขาจะมีเม็ดสีเพิ่มเติม - carotenoids. Carotenoids สามารถมองเห็นได้เฉพาะในฤดูใบไม้ร่วงเมื่อ Chlorophyl ถูกทำลายในใบริ้วรอย ฟังก์ชั่นหลักของ Chloroplasts คือการสังเคราะห์ด้วยแสง ปฏิกิริยาแสงของการไหลเวียนของการสังเคราะห์แสงบนเยื่อหุ้ม Thylacoid ซึ่งโมเลกุลคลอโรฟิลล์ได้รับการแก้ไขและปฏิกิริยาที่มืดอยู่ใน stroma ซึ่งมีเอนไซม์มากมาย

chromoplasts- เหล่านี้เป็นพลาสเดสสีเหลืองสีส้มและสีแดงที่มีเม็ดสี Carotenoid รูปร่างของ chromoplasts สามารถแตกต่างกันไปได้อย่างมีนัยสำคัญ: พวกเขาเป็นท่อทรงกลม, ผลึก ฯลฯ chromoplasts ให้ดอกไม้สีและผลไม้ของพืชดึงดูดเกสรละอองเรณูและผู้จัดจำหน่ายเมล็ดและผลไม้
leukoplasts - เหล่านี้เป็นสีขาวหรือสีฟ้าอ่อนในรอบหลักหรือ รูปไข่. พวกเขาเป็นเรื่องธรรมดาในส่วนของการตรวจจับน้ำมันของพืชเช่นในผิวหนังใบ, หัวมันฝรั่ง, ฯลฯ สารอาหารส่วนใหญ่มักจะวางแป้ง แต่พืชบางชนิดสามารถเป็นโปรตีนหรือน้ำมัน
พลาสส์จะเกิดขึ้นในเซลล์ผักจาก precipitide ซึ่งอยู่ในเซลล์ของเนื้อเยื่อเพื่อการศึกษาและมีน่องขนาดเล็กสองปู ในขั้นตอนแรกของการพัฒนา ประเภทต่าง ๆ พลาสติกมีความสามารถในการเปลี่ยนซึ่งกันและกัน: เมื่อเข้าสู่ Leukoplasts ของหัวมันฝรั่งและ chromoplasts ของรากของแครอทเป็นสีเขียว
Plasts และ Mitochondria เรียกว่า Cellides กึ่งอิสระของเซลล์เนื่องจากพวกเขามีโมเลกุล DNA ของตัวเองและ Ribosomes การสังเคราะห์โปรตีนและแบ่งออกโดยไม่คำนึงถึงการแบ่งเซลล์ คุณสมบัติเหล่านี้อธิบายโดยแหล่งกำเนิดจากสิ่งมีชีวิตแบบโปรเซสเซอร์แบบเซลล์เดียว อย่างไรก็ตาม "เอกราช" ไมโตคอนเดรียและพลาสติกมี จำกัด เนื่องจากดีเอ็นเอของพวกเขามียีนน้อยเกินไปสำหรับการดำรงอยู่ฟรีข้อมูลที่เหลือจะถูกเข้ารหัสในโครโมโซมเคอร์เนลซึ่งช่วยให้สามารถควบคุมข้อมูลของอวัยวะ
เครือข่าย endoplasmic (EPS), หรือ endoplasmic reticulum (ER)- นี่เป็นอวัยวะที่หรี่แสงได้เดี่ยวซึ่งเป็นเครือข่ายของโพรงเมมเบรนและ tubules ที่ครอบครองเนื้อหาพลาสซึมสูงถึง 30% เส้นผ่าศูนย์กลางของ eps ของช่องประมาณประมาณ 25-30 น. EPS มีสองประเภท - หยาบและราบรื่น eps หยาบ ดำเนินการไรโบโซมการสังเคราะห์โปรตีนเกิดขึ้นกับมัน eps ราบรื่นribosomes lained ฟังก์ชั่นของมันคือการสังเคราะห์ไขมันและคาร์โบไฮเดรตรวมถึงการขนส่งถุงเท้าและการทำให้เป็นกลางของสารพิษ มีการพัฒนาโดยเฉพาะอย่างยิ่งในเซลล์เหล่านั้นที่เกิดกระบวนการเผาผลาญอย่างเข้มข้นเช่นในเซลล์ของตับ - ตับ - ตับ - และเส้นใยของกล้ามเนื้อโครงร่าง สารที่สังเคราะห์ใน EPS จะถูกส่งไปยังอุปกรณ์ Golgi EPS ยังเกิดขึ้นสมัชชาเมมเบรนเซลล์ แต่การก่อตัวของพวกเขาเสร็จสมบูรณ์ในอุปกรณ์ Golgi
เครื่องจักร, golgi, หรือ คอมเพล็กซ์ Golgi- อริกะที่หรี่แสงได้เดี่ยวซึ่งเกิดขึ้นจากระบบของรถถังแบน tubules และฟองอากาศที่กินจากพวกเขา หน่วยโครงสร้างของอุปกรณ์ Golgi คือ dokyomoma - สแต็กของรถถังสำหรับหนึ่งขั้วที่สารมาจาก EPS และจากเสาตรงข้ามภายใต้การเปลี่ยนแปลงบางอย่างพวกเขาจะถูกบรรจุในฟองอากาศและถูกส่งไปยังส่วนอื่น ๆ ของเซลล์ เส้นผ่านศูนย์กลางของถังประมาณ 2 μmและฟองเล็ก ๆ ประมาณ 20-30 ไมครอน ฟังก์ชั่นหลักของ Golgji Complex - การสังเคราะห์สารบางอย่างและการปรับเปลี่ยน (เปลี่ยน) ของโปรตีนไขมันและคาร์โบไฮเดรตที่มาจาก EPS การก่อตัวครั้งสุดท้ายของเยื่อหุ้มเซลล์เช่นเดียวกับสารขนส่งในเซลล์การปรับปรุงโครงสร้างของมัน และการก่อตัวของ lysosomes ชื่อของ Golgi เป็นเกียรติของนักวิทยาศาสตร์ชาวอิตาลี Camillo Goldzhi เพื่อเป็นเกียรติแก่นักวิทยาศาสตร์ชาวอิตาลีซึ่งพบว่า organoid นี้เป็นครั้งแรก (1898)

lysosomes - มีเส้นผ่านศูนย์กลางขนาดเล็กที่หรี่แสงได้สูงถึง 1 μmซึ่งมีเอนไซม์ไฮโดรไลติกที่เกี่ยวข้องกับการย่อยเซลล์ภายในเซลล์ Membranes of Lizosom นั้นลดลงสำหรับเอนไซม์เหล่านี้ดังนั้นการดำเนินการของไลออบซ์ของฟังก์ชั่นจึงแม่นยำและกำหนดเป้าหมายมาก ดังนั้นพวกเขาจึงมีส่วนที่ใช้งานอยู่ในกระบวนการ Phagocytosis ซึ่งก่อให้เกิดฉุกเฉินทางเดินอาหารและในกรณีที่มีความอดอยากหรือความเสียหายต่อบางส่วนของเซลล์ย่อยโดยไม่ส่งผลกระทบต่ออื่น ๆ เมื่อเร็ว ๆ นี้บทบาทของ Lysosomes ในกระบวนการตายเซลล์ถูกเปิดขึ้น
vakolol - นี่คือโพรงในไซโตพลาสซึมของเซลล์และเซลล์สัตว์ จำกัด อยู่ที่เมมเบรนและเต็มไปด้วยของเหลว ในเซลล์ของสูญญากาลที่ง่ายที่สุดย่อยอาหารและหดตัวถูกตรวจพบ ครั้งแรกที่มีส่วนร่วมในกระบวนการ phagocytosis เนื่องจากพวกเขามีความแตกแยกของสารอาหาร ประการที่สองช่วยให้มั่นใจในการบำรุงรักษาสมดุลเกลือน้ำเนื่องจาก Osmoregulation ในสัตว์หลายเซลล์สูญสุญญากาศย่อยอาหารส่วนใหญ่จะพบ
ในเซลล์ผัก vacuoles จะมีอยู่เสมอพวกเขาถูกล้อมรอบด้วยเยื่อหุ้มพิเศษและเต็มไปด้วยน้ำผลไม้มือถือ เมมเบรนล้อมรอบ vacuole องค์ประกอบทางเคมีโครงสร้างและฟังก์ชั่นที่ดำเนินการอยู่ใกล้กับพลาสมาเมมเบรน น้ำผลไม้เซลลูล่า มันเป็นสารละลายที่เป็นน้ำของสารอนินทรีย์และสารอินทรีย์ต่าง ๆ รวมถึงเกลือแร่กรดอินทรีย์คาร์โบไฮเดรตโปรตีน Glycosides Alkaloids ฯลฯ สูญญากาศสามารถครอบครองได้ถึง 90% ของปริมาณของเซลล์และผลักเคอร์เนลไปที่อุปกรณ์ต่อพ่วง ส่วนนี้ของเซลล์นี้ดำเนินการถุงน่อง excretory, ออสโมติก, การป้องกัน, lysosomal และฟังก์ชั่นอื่น ๆ เนื่องจากมันสะสมสารอาหารและการเสียชีวิตมันให้ปริมาณน้ำและการบำรุงรักษารูปร่างและปริมาตรของเซลล์และยังมีเอนไซม์แยกของจำนวนมาก ส่วนประกอบของเซลล์ นอกจากนี้สารออกฤทธิ์ทางชีวภาพของ Vacuolets สามารถป้องกันไม่ให้สัตว์หลายชนิดกินพืชเหล่านี้ได้ ชุดของพืชเนื่องจากอาการบวมของฉูดฉาดเกิดขึ้นกับเซลล์ตึงเครียด
นอกจากนี้ยังมี Vacuoles ในเซลล์ของเห็ดและแบคทีเรียบางตัวอย่างไรก็ตามในเห็ดพวกเขาทำหน้าที่ของ Osmoregulation เท่านั้นและในไซยานแบคทีเรียพวกเขาสนับสนุนการลอยตัวและมีส่วนร่วมในกระบวนการของการดูดซึมของไนโตรเจนจากอากาศ
ไรโบโซม - อริโอดอยด์ขนาดเล็กที่ไม่มีขนาดเล็กที่มีเส้นผ่านศูนย์กลาง 15-20 μmประกอบด้วยสองหน่วยย่อย - ขนาดใหญ่และขนาดเล็ก Subunits Eukarot Ribosome ถูกรวบรวมในนิวเคลียวแล้วส่งไปยังไซโตพลาสซึม Ribosomes ราคา Mitochondria และ Plastids น้อยกว่า ribosome eukaryotes Subunits Ribosome รวมถึง RRNA และโปรตีน

จำนวนของ Ribosomes ในเซลล์สามารถเข้าถึงหลายสิบล้านได้หลายสิบล้าน: ในไซโตพลาสซึมไมโตคอนเดรียและพลาสต์ที่พวกเขาอยู่ในสถานะอิสระและบน eps หยาบ - ในที่เกี่ยวข้อง พวกเขามีส่วนร่วมในการสังเคราะห์โปรตีนโดยเฉพาะอย่างยิ่งดำเนินการออกอากาศ - การสังเคราะห์ทางชีวภาพของโซ่โพลีเปปไทด์บนโมเลกุลหมึก บน Ribosomes ฟรี, โปรตีน hyaloplasma, mitochondria, plastids และ ribosomes ของตัวเองถูกสังเคราะห์ในขณะที่ ribosomes ที่แนบมากับ eps หยาบ, โปรตีนจะออกอากาศเพื่อกำจัดออกจากเซลล์, การชุมนุมของเยื่อหุ้มเซลล์และ vacuoles
Ribosomes สามารถอยู่ใน Hyaloplasm หนึ่งหรือไปเป็นกลุ่มในขณะที่สังเคราะห์พร้อมกันในหนึ่ง IRNK ทันทีหลายโซ่โพลีเปปไทด์ กลุ่มดังกล่าวเรียกว่าไรโบโซม polyribosomas, หรือ polyesomas

microtubule - สิ่งเหล่านี้เป็นอวัยวะภายในกลวงกลวงแบบทรงกระบอกซึ่งซึมซับไซโตพลาสซึมทั้งหมดของเซลล์ เส้นผ่าศูนย์กลางของพวกเขาประมาณ 25 นาโนเมตรความหนาของผนังคือ 6-8 นาโนเมตร พวกเขาเกิดจากโมเลกุลโปรตีนมากมาย tubulin, รูปแบบแรกที่ 13 เธรดที่คล้ายกับลูกปัดแล้วรวบรวมใน microtubule microtubule ก่อตัวเป็นเครือข่ายไซโตพลาสซึมซึ่งให้รูปร่างและปริมาณเซลล์ผูกเมมเบรนพลาสม่ากับชิ้นส่วนอื่น ๆ ของเซลล์ให้แน่ใจว่าการขนส่งสารโดยเซลล์มีส่วนร่วมในการเคลื่อนที่ของเซลล์และส่วนประกอบของเซลล์รวมถึงใน กองวัสดุพันธุกรรม พวกเขาเป็นส่วนหนึ่งของ Cell Center และ Organoids of Motion - Flagella และ Cilia
ไมโครฟิล์ม หรือ Microniยังเป็นอวัยวะที่ไม่ลักลอบนำเข้า แต่มีรูปแบบประถมศึกษาและไม่ได้เกิดขึ้นกับ tubulin แต่ aktin. พวกเขามีส่วนร่วมในกระบวนการของการขนส่งเมมเบรนการจดจำระหว่างเซลล์การแบ่งเซลล์ไซโตพลาสซึมและในการเคลื่อนไหว ในเซลล์กล้ามเนื้อการปฏิสัมพันธ์ของ Actin Microfilaments ที่มีเกลียว Mosine ให้การลดลง
microtubule และ microfilaments สร้างโครงกระดูกเซลล์ภายใน - โครงกระดูก cytoskeleton. มันเป็นเครือข่ายที่ซับซ้อนของเส้นใยที่ให้การสนับสนุนเชิงกลสำหรับพลาสมาเมมเบรนกำหนดรูปร่างของเซลล์ตำแหน่งของช่องว่างของเซลล์และการเคลื่อนไหวของพวกเขาในระหว่างการแบ่งเซลล์

ศูนย์กลางของเซลล์ - อิงไซด์ที่ไม่ยิ้มตั้งอยู่ในเซลล์สัตว์ใกล้เคอร์เนล ในเซลล์ผักมันหายไป ความยาวประมาณ 0.2-0.3 μmและเส้นผ่าศูนย์กลาง - 0.1-0.15 μm ศูนย์เซลล์ถูกสร้างขึ้นโดยสอง centriolesนอนอยู่ในตำแหน่งร่วมกัน ระนาบตั้งฉาก, ผม. กระจ่างใสทรงกลม จาก microtubule แต่ละ centriol เกิดขึ้นจากเก้ากลุ่มของ microtubules ที่รวบรวมโดยสาม, i.e. triplets ศูนย์เซลลูล่าร์มีส่วนร่วมในกระบวนการประกอบ microtubules แบ่งวัสดุทางพันธุกรรมของเซลล์เช่นเดียวกับในการก่อตัวของ Flagella และ Cilia

ออร์แกอร์อิดเคลื่อนไหว ธง และ cilia พวกเขาเป็นเซลล์ที่ปลูกด้วย platsmable พื้นฐานของอริโอดอยด์เหล่านี้คือเก้าคู่ของ microtubules ที่ตั้งอยู่ตามขอบถนนรอบนอกและ Microtubules ฟรีสองตัวในศูนย์ microtubules เชื่อมต่อกันโดยโปรตีนต่าง ๆ ที่ให้การเบี่ยงเบนที่ตกลงกันจากแกน - การแกว่ง ความผันผวนขึ้นอยู่กับความผันผวนสูงนั่นคือพลังงานของความสัมพันธ์แบบ macroeergic ATP ถูกใช้ไปกับกระบวนการนี้ การฟื้นฟูรสชาติที่หายไปและ Cilia เป็นฟังก์ชั่น ฐานราศีพฤษภ, หรือ kinetosomตั้งอยู่ในรากฐานของพวกเขา
ความยาวปรับเลนส์ประมาณ 10-15 นาโนเมตรและ Flagellas เป็น 20-50 ไมครอน เนื่องจากการเคลื่อนที่อย่างเคร่งครัดของ Flagella และ ciliations ไม่เพียง แต่การเคลื่อนไหวของสัตว์เซลล์เดียวอสุจิ ฯลฯ แต่ยังเกิดขึ้นเพื่อทำความสะอาดระบบทางเดินหายใจการส่งเสริมไข่ในท่อมดลูกเนื่องจากชิ้นส่วนเหล่านี้ทั้งหมดของ ร่างกายมนุษย์ถูกล่อลวงด้วยเยื่อบุผิวลูกเรือ

การรวม
การรวม- เหล่านี้เป็นส่วนประกอบของเซลล์ที่ไม่ถาวรที่เกิดขึ้นและหายไปในกระบวนการทำมาหากิน สิ่งเหล่านี้มีทั้งสารอะไหล่เช่นแป้งแป้งหรือโปรตีนในเซลล์พืชเม็ดไกลโคเจนในเซลล์สัตว์และเห็ดที่ลอยตัวหยดไขมันในเซลล์และของเสียทุกชนิดโดยเฉพาะอย่างยิ่งการแก้ไขผลกระทบที่เป็นผลมาจากการก่อตัวของ phagocytosis น่อง

ความสัมพันธ์ของโครงสร้างและหน้าที่ของชิ้นส่วนและเซลล์อิงเกณฑ์ - พื้นฐานของความซื่อสัตย์
แต่ละชิ้นส่วนของเซลล์ในมือข้างหนึ่งเป็นโครงสร้างที่แยกต่างหากที่มีโครงสร้างและฟังก์ชั่นที่เฉพาะเจาะจงและอื่น ๆ ส่วนประกอบของระบบที่ซับซ้อนมากขึ้นที่เรียกว่าเซลล์ ส่วนที่ใหญ่ที่สุดของข้อมูลทางพันธุกรรมของเซลล์ยูคาริโอตนั้นมีความเข้มข้นในแกนกลาง แต่เคอร์เนลนั้นไม่สามารถให้การดำเนินการได้เนื่องจากสิ่งนี้จำเป็นสำหรับอย่างน้อยไซโตพลาสซึมทำหน้าที่เป็นสารพื้นฐานและไรโบโซม ซึ่งการสังเคราะห์นี้เกิดขึ้น ไรโบโซมส่วนใหญ่ตั้งอยู่บนเครือข่าย endoplasmic เม็ดจากที่โปรตีนส่วนใหญ่มักส่งไปยังคอมเพล็กซ์ Golgi แล้วหลังจากการดัดแปลง - ในส่วนเหล่านั้นของเซลล์ที่พวกเขามีจุดประสงค์หรือออกไปด้านนอก บรรจุภัณฑ์เมมเบรนของโปรตีนและคาร์โบไฮเดรตสามารถรวมเข้ากับเมมเบรนอิงอริกและเมมเบรนพลาสซึมให้การอัพเดทอย่างต่อเนื่อง คอมเพล็กซ์ Golgji ยังเต็มไปด้วยฟังก์ชั่นที่สำคัญที่สุดของ lysosomes และ vacuoles ตัวอย่างเช่นไม่มี lysosomes เซลล์จะกลายเป็นรูปของโมเลกุลและโครงสร้างของเสียอย่างรวดเร็ว
การไหลของกระบวนการทั้งหมดเหล่านี้ต้องการพลังงานที่ผลิตโดยไมโตคอนเดรียและในพืช - และคลอโรพลาสต์ และถึงแม้ว่าอริโอดอยด์เหล่านี้จะค่อนข้างเป็นอิสระเพราะพวกเขามีโมเลกุลของดีเอ็นเอของตัวเองโปรตีนบางอย่างของพวกเขายังคงถูกเข้ารหัสโดยจีโนมนิวเคลียร์และสังเคราะห์ในไซโตพลาสซึม
ดังนั้นเซลล์จึงเป็นเอกภาพที่แยกกันไม่ออกของส่วนประกอบของส่วนประกอบแต่ละอย่างจะทำหน้าที่เฉพาะของตัวเอง
การเผาผลาญและการเปลี่ยนแปลงของพลังงานคือคุณสมบัติของสิ่งมีชีวิต การแลกเปลี่ยนพลังงานและพลาสติกความสัมพันธ์ของพวกเขา ขั้นตอนการแลกเปลี่ยนพลังงาน การหมักและการหายใจ การสังเคราะห์ด้วยแสงความหมายของมันบทบาทจักรวาล ขั้นตอนการสังเคราะห์แสง ปฏิกิริยาการสังเคราะห์แสงที่เบาและมืดความสัมพันธ์ของพวกเขา Chemosynthesis บทบาทของแบคทีเรียเคมีสังเคราะห์บนโลก
การเผาผลาญและการเปลี่ยนแปลงพลังงาน - คุณสมบัติของสิ่งมีชีวิต
เซลล์สามารถเป็นโรงงานเคมีขนาดเล็กซึ่งเกิดขึ้นหลายร้อยและนับพันของปฏิกิริยาเคมี
การเผาผลาญ - การรวมกันของการเปลี่ยนแปลงทางเคมีที่มีวัตถุประสงค์เพื่อรักษาและการเล่นระบบชีวภาพในตัวเอง
มันรวมถึงการเข้ารับสารในร่างกายในกระบวนการของโภชนาการและการหายใจการเผาผลาญในเซลล์หรือ การเผาผลาญรวมถึงการจัดสรรผลิตภัณฑ์ Exchange Finite
การเผาผลาญมีการเชื่อมโยงอย่างแยกไม่ออกกับการเปลี่ยนแปลงของพลังงานหนึ่งประเภทให้กับผู้อื่น ตัวอย่างเช่นในกระบวนการของการสังเคราะห์ด้วยแสงพลังงานแสงได้รับการคุ้มครองในรูปแบบของพลังงานของพันธะเคมีของโมเลกุลอินทรีย์ที่ซับซ้อนและในกระบวนการของการหายใจมันถูกปล่อยออกมาและใช้เวลาในการสังเคราะห์โมเลกุลใหม่งานกลไกและออสโมติก กระจายไปเป็นความร้อนและอื่น ๆ
การไหลของปฏิกิริยาเคมีในสิ่งมีชีวิตที่มีชีวิตมั่นใจได้โดยตัวเร่งปฏิกิริยาทางชีวภาพของธรรมชาติโปรตีน - เอนไซม์, หรือ เอนไซม์. เช่นเดียวกับตัวเร่งปฏิกิริยาอื่น ๆ เอนไซม์ช่วยเร่งการไหลของปฏิกิริยาเคมีในเซลล์ในสิบและหลายแสนเท่าและบางครั้งพวกเขาทำ แต่ไม่เปลี่ยนแปลงธรรมชาติหรือคุณสมบัติของผลิตภัณฑ์ขั้นสุดท้าย (ผลิตภัณฑ์) ของปฏิกิริยาและ อย่าเปลี่ยนตัวเอง เอนไซม์สามารถเป็นทั้งโปรตีนที่เรียบง่ายและซับซ้อนซึ่งนอกเหนือไปจากส่วนโปรตีนยังเป็นโปรตีน - cofactor (Coenzyme). ตัวอย่างของเอนไซมม์คือน้ำลายอะไมเลส, การแยกโพลีแซคคาไรด์ที่มีการเคี้ยวเป็นเวลานานและเป๊ปซินให้การย่อยของโปรตีนในกระเพาะอาหาร
เอนไซม์แตกต่างจากตัวเร่งปฏิกิริยาธรรมชาติที่ไม่ใช่โลหะที่มีความจำเพาะสูงเพิ่มขึ้นอย่างมีนัยสำคัญกับความช่วยเหลือของอัตราการเกิดปฏิกิริยารวมถึงความเป็นไปได้ที่จะควบคุมการกระทำโดยการเปลี่ยนเงื่อนไขของปฏิกิริยาหรือโต้ตอบกับพวกเขาจากสารต่าง ๆ นอกจากนี้เงื่อนไขที่มีการเร่งปฏิกิริยาของเอนไซม์แตกต่างกันอย่างมีนัยสำคัญจากผู้ที่อยู่ภายใต้ Neopenmen: อุณหภูมิที่เหมาะสมที่สุดสำหรับการทำงานของเอนไซม์ในร่างกายมนุษย์คืออุณหภูมิของ $ 37 ° C $ ความดันจะต้องใกล้เคียง บรรยากาศและสภาพแวดล้อม $ RAL $ สามารถลังเลอย่างมีนัยสำคัญ ดังนั้นสำหรับอะไมเลสสื่ออัลคาไลน์เป็นสิ่งจำเป็นและสำหรับเป๊ปซีน - เปรี้ยว
กลไกของการกระทำของเอนไซม์คือการลดพลังงานกระตุ้นการเปิดใช้งานของสาร (พื้นผิว) เนื่องจากการก่อตัวของคอมเพล็กซ์ของเอนไซม์ระดับกลาง

การแลกเปลี่ยนพลังงานและพลาสติกความสัมพันธ์ของพวกเขา
การเผาผลาญประกอบด้วยสองกระบวนการพร้อมกันในเซลล์: การแลกเปลี่ยนพลาสติกและพลังงาน
แลกเปลี่ยนพลาสติก (Anabolism, การดูดกลืน) มันเป็นการผสมผสานระหว่างปฏิกิริยาการสังเคราะห์ที่ไปกับต้นทุนของพลังงาน ATP ในกระบวนการของการเผาผลาญพลาสติกสารอินทรีย์ที่เซลล์ที่เซลล์ต้องการสังเคราะห์ ตัวอย่างของปฏิกิริยาการแลกเปลี่ยนพลาสติกคือการสังเคราะห์ด้วยแสงการสังเคราะห์โปรตีนและการจำลองแบบ (การตัดเย็บด้วยตนเอง) DNA
การแลกเปลี่ยนพลังงาน (catabolism, การกระจาย) - นี่คือการรวมกันของปฏิกิริยาของสารแยกที่ซับซ้อนเพื่อที่ง่ายกว่า อันเป็นผลมาจากการแลกเปลี่ยนพลังงานพลังงานจะถูกเน้นในรูปแบบของ ATP กระบวนการที่สำคัญที่สุดของการแลกเปลี่ยนพลังงานคือการหายใจและการหมัก
การแลกเปลี่ยนพลาสติกและพลังงานมีการเชื่อมโยงอย่างแยกไม่ออกเนื่องจากสารอินทรีย์ถูกสังเคราะห์ในระหว่างการแลกเปลี่ยนพลาสติกและพลังงานของ ATP เป็นสิ่งจำเป็นและในกระบวนการของการแลกเปลี่ยนพลังงานสารอินทรีย์จะถูกแยกและพลังงานออกมาซึ่งจะถูกใช้ไป กระบวนการสังเคราะห์
พลังงานของสิ่งมีชีวิตจะได้รับในกระบวนการโภชนาการและพวกเขาปล่อยมันและแปลเป็นรูปแบบราคาไม่แพงในระหว่างกระบวนการทางเดินหายใจ โดยวิธีการของโภชนาการสิ่งมีชีวิตทั้งหมดจะถูกแบ่งออกเป็น Autotrophs และ Heterotrophs aptotropy สามารถสังเคราะห์สารอินทรีย์ได้อย่างอิสระจากอนินทรีย์และ heterotrophs ใช้สารอินทรีย์สำเร็จรูปแบบครบครัน
ขั้นตอนของการแลกเปลี่ยนพลังงาน
แม้จะมีความซับซ้อนของปฏิกิริยาการแลกเปลี่ยนพลังงาน แต่ก็แบ่งตามอัตภาพเป็นสามขั้นตอน: การเตรียมการแบบไม่ใช้ออกซิเจน (ออกซิเจน) และแอโรบิก (ออกซิเจน)
บน เวทีเตรียมความพร้อม โมเลกุลของโพลีแซคคาไรด์, ไขมัน, โปรตีน, กรดนิวกิคสลายตัวเพื่อง่ายขึ้นเช่นกลูโคสกลีเซอรีนและกรดไขมันกรดอะมิโนนิวคลีโอไทด์ ฯลฯ ขั้นตอนนี้สามารถเกิดขึ้นได้โดยตรงในเซลล์หรือในลำไส้จากที่มีการส่งสารแยก ด้วยการไหลเวียนของเลือด
แบบไม่ใช้ออกซิเจน การแลกเปลี่ยนพลังงานมาพร้อมกับการแยกโมโนเมอร์สารประกอบอินทรีย์ต่อไปเพื่อผลิตภัณฑ์ระดับกลางที่ง่ายกว่าเช่นกรด pyruogradic หรือ pyruvate มันไม่จำเป็นต้องมีออกซิเจนและสำหรับสิ่งมีชีวิตจำนวนมากที่อาศัยอยู่ใน Silt Swamp หรือในลำไส้ของบุคคลเป็นวิธีเดียวที่จะผลิตพลังงาน ขั้นตอนการแลกเปลี่ยนพลังงานแบบไม่ใช้ออกซิเจนในไซโตพลาสซึม
สารที่แตกต่างกันอาจอยู่ภายใต้ความแตกแยกออกซิเจน แต่มันค่อนข้างบ่อยครั้งที่พื้นผิวกลูโคส เรียกว่ากระบวนการแยกออกจากวัว คนโง่. ด้วยโมเลกุลกลูโคสโมเลกุลกลูโคสสูญเสียสี่อะตอมไฮโดรเจน i.e. มันเป็นออกซิไดซ์โมเลกุลสองของกรดนกยูงจะเกิดขึ้นโมเลกุล ATP สองตัวและสองโมเลกุลของผู้ให้บริการไฮโดรเจนที่ลดลง $ NAPN + H ^ (+) $:
$ с_6н_ (12) O_6 + 2N_3RO_4 + 2ADF + 2NV → 2S_3N_4O_3 + 2AF + 2NV + H ^ (+) + 2N_2O $
การก่อตัวของ ATP จาก ADP เกิดจากการถ่ายโอนโดยตรงของประจุลบฟอสเฟตด้วยน้ำตาลก่อน phosphorylated และเรียกว่า สารตั้งต้น phosphorylation
เวทีแอโรบิก การแลกเปลี่ยนพลังงานสามารถเกิดขึ้นได้เฉพาะในการปรากฏตัวของออกซิเจนในขณะที่สารประกอบระดับกลางเกิดขึ้นในกระบวนการแยกออกซิเจนออกซิไดซ์เพื่อผลิตภัณฑ์ขั้นสุดท้าย (คาร์บอนไดออกไซด์และน้ำ) และพลังงานส่วนใหญ่ที่เก็บไว้ในพันธะเคมีของสารประกอบอินทรีย์ที่โดดเด่น มันเข้าสู่พลังงานของ Macroeergic Ties 36 โมเลกุล ATP ขั้นตอนนี้เรียกอีกอย่างว่า การหายใจผ้า. ในกรณีที่ไม่มีออกซิเจนตัวกลางจะถูกแปลงเป็นสารอินทรีย์อื่น ๆ และกระบวนการนี้เรียกว่า การหมัก
ลมหายใจ
กลไกของการหายใจของเซลล์จะแสดงเป็นแผนผังในรูปที่

การหายใจแบบแอโรบิคเกิดขึ้นในไมโตคอนเดรียในขณะที่กรดแบบพีโรคอกรสูญเสียอะตอมคาร์บอนหนึ่งอะตอมซึ่งมาพร้อมกับการสังเคราะห์หนึ่งการลดการเทียบเท่า $ nad / h ^ (+) $ และ acetylcoferment โมเลกุล A (Acetyl-Coa):
$ с_3н_4o_3 + nad + h ~ coa → ch_3co ~ coa + nadn + h ^ (+) + co_2 $
Acetyl-Coa ใน Matrix Mitochondria มีส่วนร่วมในสายโซ่ของปฏิกิริยาเคมีจำนวนทั้งที่เรียกว่า crec cycle (วัฏจักรของกรด Tricarboxylic วงจรกรดซิตริก. ในระหว่างการเปลี่ยนแปลงเหล่านี้โมเลกุล ATP สองตัวเกิดขึ้น Acetyl-COA นั้นออกซิไดซ์เป็นก๊าซคาร์บอนไดออกไซด์อย่างสมบูรณ์และไอออนไฮโดรเจนและอิเล็กตรอนจะติดอยู่กับผู้ให้บริการไฮโดรเจน $ na / n ^ (+) $ และ $ fadn_2 $ ผู้ให้บริการขนส่งโปรตอนไฮโดรเจนและอิเล็กตรอนไปยังเยื่อหุ้มภายในของการขึ้นรูปไมโตคอนเดรีย ด้วยความช่วยเหลือของโปรตีนของผู้ให้บริการ, โปรตีนไฮโดรเจนจะถูกฉีดเข้าไปในอวกาศ intermogram และอิเล็กตรอนจะถูกส่งไปตามห่วงโซ่ระบบทางเดินหายใจของเอนไซม์ที่ตั้งอยู่บนเมมเบรนภายในของไมโตคอนเดรียและจะถูกรีเซ็ตเป็นอะตอมออกซิเจน:
$ o_2 + 2e ^ (-) → o_2 ^ - $
ควรสังเกตว่าโปรตีนโซ่ระบบทางเดินหายใจบางชนิดมีธาตุเหล็กและซัลเฟอร์
จากพื้นที่ intermambrane โปรตอนไฮโดรเจนจะถูกส่งกลับไปสู่ \u200b\u200bMatrix Mitochondria โดยใช้เอนไซม์พิเศษ - ATP-Synthase และพลังงานที่เผยแพร่ในการสังเคราะห์โมเลกุล ATP 34 โมเลกุลจากโมเลกุลกลูโคสแต่ละโมเลกุล กระบวนการนี้เรียกว่า phosphorylation ออกซิเดชัน. ใน Matrix Mitochondria โปรตอนไฮโดรเจนตอบโต้ด้วยอนุมูลของออกซิเจนด้วยการก่อตัวของน้ำ:
$ 4H ^ (+) + o_2 ^ - → 2H_2O $
การรวมกันของปฏิกิริยาการหายใจออกซิเจนสามารถแสดงได้ดังนี้:
$ 2с_3н_4O_3 + 6O_2 + 36NNN_3RO_4 + 36ADF → 6SO_2 + 38N_2O + 36ADF. $
สมการทางเดินหายใจทั้งหมดมีลักษณะดังนี้:
$ S_6N_ (12) O_6 + 6O_2 + 38N_3RO_4 + 38ADF → 6SO_2 + 40N_2O + 38ATF. $
การหมัก
ในกรณีที่ไม่มีออกซิเจนหรือขาดการหมักเกิดขึ้น การทอดเป็นวิธีการวิวัฒนาการก่อนหน้านี้ของการได้รับพลังงานมากกว่าการหายใจอย่างไรก็ตามมันมีผลกำไรน้อยลงเนื่องจากสารอินทรีย์เกิดขึ้นเนื่องจากการหมักยังคงเป็นพลังงานที่อุดมไปด้วย มีการหมักพื้นฐานหลายประเภท: กรดแลคติค, แอลกอฮอล์, กรดอะซิติก ฯลฯ ดังนั้นในกล้ามเนื้อโครงร่างในกรณีที่ไม่มีออกซิเจนในระหว่างการหมัก, กรดเกรดเพียร์จะได้รับการฟื้นฟูเป็นกรดแลคติกในขณะที่การกู้คืนที่เกิดขึ้นก่อนหน้านี้ และอีกสองโมเลกุล ATP ยังคงอยู่:
$ 2с_3н_4O_3 + 2NADN + H ^ (+) → 2C_3N_6O_3 + 2ND $
เมื่อหมักกับเห็ดยีสต์กรดเกรดเพียร์ในการปรากฏตัวของออกซิเจนกลายเป็นเอทิลแอลกอฮอล์และคาร์บอนมอนอกไซด์ (IV):
$ с_3н_4O_3 + NAPH + H ^ (+) → c_2n_5on + co_2 + มากกว่า ^ (+) $
ในกรณีที่มีการหมักด้วยความช่วยเหลือของจุลินทรีย์จากกรดเกรดเพียร์อะซิติกน้ำมันกรดฟอร์มิก ฯลฯ ก็สามารถเกิดขึ้นได้
ATP ที่ได้รับเป็นผลมาจากการแลกเปลี่ยนพลังงานถูกบริโภคในเซลล์ในงานประเภทต่าง ๆ : เคมี, ออสโมติก, ไฟฟ้า, เครื่องกลและกฎระเบียบ งานเคมีอยู่ที่การสังเคราะห์จากโปรตีนไขมันคาร์โบไฮเดรตกรดนิวคลีอิกและสารประกอบสำคัญอื่น ๆ งานออสโมติกรวมถึงกระบวนการดูดซึมของเซลล์และการลบสารจากมันซึ่งในพื้นที่นอกเซลล์มีความเข้มข้นมีขนาดใหญ่กว่าในเซลล์เอง งานไฟฟ้าเชื่อมต่อกันอย่างใกล้ชิดกับออสโมติกเนื่องจากเป็นผลมาจากการเคลื่อนย้ายอนุภาคที่มีประจุผ่านเมมเบรนการเรียกเก็บเงินเมมเบรนจะเกิดขึ้นและคุณสมบัติของความตื่นเต้นเร้าความสามารถและการนำไฟฟ้า งานเครื่องกลคือคอนจูเกตกับการเคลื่อนไหวของสารและโครงสร้างภายในเซลล์เช่นเดียวกับเซลล์ทั่วไป งานกำกับดูแลรวมถึงกระบวนการทั้งหมดที่มีวัตถุประสงค์เพื่อประสานงานกระบวนการในเซลล์
การสังเคราะห์ด้วยแสง, ความหมาย, บทบาทอวกาศ
การสังเคราะห์แสง เรียกกระบวนการเปลี่ยนพลังงานของแสงเป็นพลังงานของพันธะเคมีของสารประกอบอินทรีย์กับการมีส่วนร่วมของคลอโรฟิลล์
อันเป็นผลมาจากการสังเคราะห์ด้วยแสงประมาณ 150 พันล้านตันของสารอินทรีย์และออกซิเจนประมาณ 200 พันล้านตันเป็นประจำทุกปี กระบวนการนี้ให้วงจรคาร์บอนในชีวมณฑลโดยไม่ให้การสะสมกับก๊าซคาร์บอนไดออกไซด์และป้องกันการเกิดขึ้นของผลเรือนกระจกและความร้อนสูงเกินไปของโลก สารอินทรีย์ที่เกิดจากการสังเคราะห์ด้วยแสงที่ไม่มีสิ่งมีชีวิตอื่นอย่างสมบูรณ์เป็นส่วนสำคัญของพวกเขาสำหรับหลายล้านปีได้ก่อตัวเป็นเงินฝากของแร่ธาตุ (หินและถ่านหินสีน้ำตาลน้ำมัน) เมื่อเร็ว ๆ นี้น้ำมันเรพซีด ("ไบโอดีเซล") และแอลกอฮอล์ที่ได้รับจากสิ่งตกค้างพืชเริ่มใช้เป็นเชื้อเพลิง โอโซนฟอร์มออกซิเจนภายใต้การกระทำของการปล่อยไฟฟ้าซึ่งเป็นหน้าจอโอโซนที่ปกป้องทุกอย่างที่มีชีวิตอยู่บนโลกจากผลการทำลายล้างของรังสีอัลตราไวโอเลต
เพื่อนร่วมชาติของเรานักสรีรวิทยาที่โดดเด่นของพืช K. A. Timiryazev (1843-1920) เรียกว่าบทบาทของการสังเคราะห์ด้วยแสงของ "จักรวาล" ในขณะที่เขาผูกดินแดนกับดวงอาทิตย์ (อวกาศ) ให้การไหลเข้าของพลังงานบนโลก
ขั้นตอนการสังเคราะห์แสง ปฏิกิริยาการสังเคราะห์แสงที่มืดและมืดความสัมพันธ์ของพวกเขา
ในปี 1905 นักสรีรวิทยาชาวอังกฤษของพืช F. แบล็กแมนพบว่าความเร็วของการสังเคราะห์ด้วยแสงไม่สามารถเพิ่มขึ้นไม่เกี่ยวข้องปัจจัยบางอย่าง จำกัด ขึ้นอยู่กับสิ่งนี้เขาหยิบยกสมมติฐานของการปรากฏตัวของสองขั้นตอนของการสังเคราะห์ด้วยแสง: เบา และ temnova. ด้วยความเข้มของแสงน้อยความเร็วของปฏิกิริยาแสงเพิ่มขึ้นในสัดส่วนการเพิ่มขึ้นของพลังของแสงและนอกจากนี้ปฏิกิริยาเหล่านี้ไม่ได้ขึ้นอยู่กับอุณหภูมิเนื่องจากพวกเขาไม่ต้องการเอนไซม์ ปฏิกิริยาแสงดำเนินต่อไปในเยื่อหุ้ม Tylacoid
อัตราของปฏิกิริยาที่มืดในทางตรงกันข้ามเพิ่มขึ้นตามอุณหภูมิที่เพิ่มขึ้น แต่เมื่อถึงขีด จำกัด อุณหภูมิที่ $ 30 ° C $ การเติบโตนี้ถูกยกเลิกซึ่งบ่งชี้ลักษณะของเอนไซม์ของการเปลี่ยนแปลงเหล่านี้ที่เกิดขึ้นใน stroma ควรสังเกตว่าแสงยังมีอิทธิพลบางอย่างต่อปฏิกิริยาสีเข้มแม้จะมีความจริงที่ว่าพวกเขาเรียกว่ามืด

ขั้นตอนแสงของการสังเคราะห์แสงที่ดำเนินการบนเยื่อหุ้ม tylakoid ที่ดำเนินการคอมเพล็กซ์โปรตีนหลายประเภทหลัก ๆ ซึ่งเป็นระบบภาพถ่าย I และ II รวมถึง ATFSINTASE ระบบถ่ายภาพรวมถึงคอมเพล็กซ์เม็ดสีที่มี carotenoids นอกจากนี้ยังมีคลอโรฟิลล์ แคโรทีนอยด์จับแสงในพื้นที่สเปกตรัมที่ไม่ได้คลอโรฟิลล์เช่นเดียวกับปกป้องคลอโรฟิลล์จากการทำลายด้วยแสงที่มีความเข้มสูง
นอกเหนือจากคอมเพล็กซ์เม็ดสีแล้วระบบ Photosys ยังมีโปรตีนตัวรับอิเล็กตรอนจำนวนหนึ่งที่ส่งอิเล็กตรอนซึ่งกันและกันอย่างต่อเนื่องจากโมเลกุลคลอโรฟิลล์ ลำดับของโปรตีนเหล่านี้เรียกว่า วงจรไฟฟ้าของคลอโรพลาสต์
ด้วย Photo System II คอมเพล็กซ์โปรตีนพิเศษยังมีความเกี่ยวข้องเช่นกันซึ่งช่วยให้มั่นใจว่าออกซิเจนในกระบวนการสังเคราะห์แสง คอมเพล็กซ์ออกซิเจนนี้มีไอออนแมงกานีสและคลอรีน
ใน เฟสแสง แสง Quanta หรือโฟตอนที่เข้าสู่โมเลกุลคลอโรฟิลล์ตั้งอยู่บนเยื่อหุ้ม Tylacoid แปลพวกเขาเป็นรัฐที่ตื่นเต้นที่โดดเด่นด้วยพลังงานอิเล็กตรอนที่สูงขึ้น ในกรณีนี้อิเล็กตรอนที่ตื่นเต้นจากระบบถ่ายภาพคลอโรฟิลล์ฉันจะถูกส่งผ่านห่วงโซ่ตัวกลางไปยังผู้ให้บริการไฮโดรเจนของ NADF ซึ่งเชื่อมต่อโปรตอนของไฮโดรเจนมีให้บริการในสารละลายที่เป็นน้ำเสมอ:
$ NADF + 2E ^ (-) + 2H ^ (+) → napfn + h ^ (+) $
กู้คืน $ napfn + h ^ (+) $ จะถูกใช้ในภายหลังในเวทีมืด อิเล็กตรอนจาก Chlorophyll Photosystems II จะถูกส่งไปตามหลุมอิเล็กทรอนิกส์พวกเขาเติมเต็มหลุมอิเล็กทรอนิกส์ของ Chlorophyll Photosystems I. การขาดอิเล็กตรอนใน Chlorophyll Photosystems II เต็มเนื่องจากไม่แน่นอนในโมเลกุลของน้ำซึ่งเกิดขึ้นกับการมีส่วนร่วมของการเข้าร่วม สารประกอบที่กล่าวถึง อันเป็นผลมาจากการสลายตัวของโมเลกุลของน้ำซึ่งเรียกว่า photolisom, โปรตอนไฮโดรเจนจะเกิดขึ้นและออกซิเจนโมเลกุลมีความโดดเด่นซึ่งเป็นผลพลอยได้จากการสังเคราะห์ด้วยแสง:
$ h_2o → 2H ^ (+) + 2e ^ (-) + (1) / (2) o_2 $
ข้อมูลทางพันธุกรรมในเซลล์ ยีนรหัสพันธุกรรมและคุณสมบัติของมัน ลักษณะของเมทริกซ์ธรรมชาติของปฏิกิริยาการสังเคราะห์การสังเคราะห์ Biosynthesis ของโปรตีนและกรดนิวคลีอิก
ข้อมูลทางพันธุกรรมในเซลล์
การเล่นตัวเองเช่นนี้เป็นหนึ่งในคุณสมบัติพื้นฐานของการใช้ชีวิต เนื่องจากปรากฏการณ์นี้มีความคล้ายคลึงกันไม่เพียง แต่ระหว่างสิ่งมีชีวิต แต่ระหว่างเซลล์แต่ละเซลล์เช่นเดียวกับอริกไมออน (ไมโตคอนเดรียและพลาสต์) พื้นฐานวัสดุของความคล้ายคลึงกันนี้คือการถ่ายโอนข้อมูลทางพันธุกรรมที่เข้ารหัสในลำดับของนิวคลีโอไทด์ซึ่งดำเนินการเนื่องจากกระบวนการของการจำลองแบบ (ความนับถือตนเอง) DNA คุณสมบัติและคุณสมบัติทั้งหมดของเซลล์และสิ่งมีชีวิตเนื่องจากโปรตีนซึ่งมีโครงสร้างส่วนใหญ่กำหนดโดย DNA Nucleotide Sequences ดังนั้นการสังเคราะห์ทางชีวภาพของกรดนิวคลีอิกและโปรตีนจึงมีความสำคัญอย่างยิ่งในกระบวนการเผาผลาญ หน่วยโครงสร้างของข้อมูลทางพันธุกรรมคือยีน
ยีนรหัสพันธุกรรมและคุณสมบัติของมัน
ข้อมูลทางพันธุกรรมในกรงไม่ได้อยู่ในเสาหินมันแบ่งออกเป็น "คำ" แยกต่างหาก - ยีน
ยีน - นี่คือหน่วยพื้นฐานของข้อมูลทางพันธุกรรม
ทำงานในโครงการยีนของมนุษย์ซึ่งจัดขึ้นพร้อมกันในหลายประเทศและเสร็จสิ้นในช่วงต้นศตวรรษนี้ทำให้เราเข้าใจว่าบุคคลมีเพียงประมาณ 25-30,000 ยีน แต่ข้อมูลจาก DNA ส่วนใหญ่ของเราไม่เคยอ่าน เนื่องจากมีไซต์ที่ไม่มีความหมายจำนวนมากซ้ำและการเข้ารหัสยีนที่สูญเสียความสำคัญสำหรับบุคคล (หางร่างกายเจ็บ ฯลฯ ) นอกจากนี้ยังมีการถอดรหัสหลายยีนที่รับผิดชอบในการพัฒนาโรคทางพันธุกรรมเช่นเดียวกับยีนเป้าหมายของยาเสพติด อย่างไรก็ตามการประยุกต์ใช้งานจริงของผลลัพธ์ที่ได้รับในระหว่างการดำเนินการของโปรแกรมนี้จะถูกเลื่อนออกไปจนกว่ายีนของผู้คนจำนวนมากจะถูกถอดรหัสและกลายเป็นที่ชัดเจนว่าสิ่งที่พวกเขายังคงแตกต่างกัน
ยีนที่เข้ารหัสโครงสร้างหลักของโปรตีน Ribosomal หรือ RNA ขนส่งเรียกว่า เกี่ยวกับโครงสร้างและยีนที่ให้การเปิดใช้งานหรือระงับการอ่านข้อมูลจากยีนโครงสร้าง - เกี่ยวกับการกำกับดูแล. อย่างไรก็ตามแม้แต่ยีนโครงสร้างยังมีส่วนการกำกับดูแล
ข้อมูลทางพันธุกรรมของสิ่งมีชีวิตถูกเข้ารหัสเป็น DNA ในรูปแบบของการรวมกันบางอย่างของนิวคลีโอไทด์และลำดับของพวกเขา - รหัสพันธุกรรม. คุณสมบัติของมันคือ: Triplet, ความจำเพาะ, ความเก่งกาจ, ความซ้ำซ้อนและการไม่สามารถ reveanybility นอกจากนี้ยังไม่มีเครื่องหมายวรรคตอนในรหัสพันธุกรรม
กรดอะมิโนแต่ละตัวถูกเข้ารหัสใน DNA สามนิวคลีโอไทด์ - แฝด ตัวอย่างเช่น Methionine ถูกเข้ารหัสโดย TAC Triplet นั่นคือรหัสของ Triplet ในทางกลับกันแต่ละแฝดจะเข้ารหัสเพียงกรดอะมิโนเดียวซึ่งเป็นความจำเพาะหรือความกระจ่างใส รหัสพันธุกรรมเป็นสากลสำหรับสิ่งมีชีวิตทั้งหมดนั่นคือข้อมูลทางพันธุกรรมเกี่ยวกับโปรตีนของมนุษย์สามารถพิจารณาแบคทีเรียและในทางกลับกัน สิ่งนี้บ่งบอกถึงความสามัคคีของต้นกำเนิดของโลกอินทรีย์ อย่างไรก็ตามการรวมกัน 64 ชุดของนิวคลีโอไทด์ที่สามสอดคล้องกับกรดอะมิโนเพียง 20 กรดซึ่งเป็นผลมาจากกรดอะมิโนหนึ่งที่สามารถเข้ารหัส 2-6 ทริปิลนั่นคือรหัสพันธุกรรมที่มากเกินไปหรือเสื่อมโทรม สาม Triplets ไม่มีกรดอะมิโนที่สอดคล้องกันพวกเขาเรียกว่า หยุด codonsเนื่องจากพวกเขาแสดงถึงจุดสิ้นสุดของการสังเคราะห์โซ่โพลีเปปไทด์
ลำดับพื้นฐานใน DNA Triplets และกรดอะมิโนที่เข้ารหัสโดยพวกเขา

* หยุด Codon ซึ่งหมายถึงจุดสิ้นสุดของการสังเคราะห์โซ่โพลีเปปไทด์
การลดชื่อของกรดอะมิโน:
Ala - Alanine
arg - อาร์จินีน
ASN - Asparagin
ASP - Asparagic Acid
วาล - วาลิน
GIS - GISTIDIN
gly - glycine
Gln - กลูตามีน
ความลึก - กรดกลูตามิก
Ile - Isoleucine
lei - leucin
ลิซ - ไลซีน
meti - methionine
Pro - proline
อ่อนโยน
tir - tyrosine
Tre - Treonin
สาม - ทริปโตเฟน
เครื่องเป่าผม - Phenylalanine
CIS - Cysteine
หากคุณเริ่มอ่านข้อมูลทางพันธุกรรมที่ไม่ได้มาจากนิวคลีโอไทด์แรกใน Triplet แต่จากที่สองไม่เพียง แต่การเปลี่ยนเฟรมการอ่านจะเกิดขึ้น - โปรตีนสังเคราะห์ในลักษณะนี้จะแตกต่างกันอย่างสมบูรณ์ไม่เพียง แต่ในลำดับนิวคลีโอไทด์ แต่ยังรวมถึง โครงสร้างและคุณสมบัติ ไม่มีสัญญาณเครื่องหมายวรรคตอนระหว่าง Triplets ดังนั้นจึงไม่มีอุปสรรคต่อกรอบการอ่านซึ่งเปิดพื้นที่สำหรับการเกิดขึ้นและการบันทึกการกลายพันธุ์
ธรรมชาติของปฏิกิริยาการสังเคราะห์ทางชีวภาพ
เซลล์แบคทีเรียสามารถเพิ่มเป็นสองเท่าทุก ๆ 20-30 นาทีและ Eukaryotes ทุกวันและบ่อยครั้งยิ่งขึ้นซึ่งต้องใช้ความเร็วสูงและแม่นยำของการจำลองแบบ DNA นอกจากนี้แต่ละเซลล์มีสำเนาของโปรตีนหลายร้อยและหลายพันรายการโดยเฉพาะเอนไซม์ดังนั้นในการเล่นวิธีการผลิต "ชิ้น" ที่ยอมรับไม่ได้ของการผลิต วิธีการที่ก้าวหน้ามากขึ้นคือการปั๊มซึ่งช่วยให้คุณได้รับสำเนาที่แม่นยำของผลิตภัณฑ์จำนวนมากและนอกจากนี้ยังลดต้นทุน สำหรับการปั๊มเมทริกซ์จำเป็นต้องมีการแสดงความประทับใจ
ในเซลล์หลักการสังเคราะห์เมทริกซ์คือโปรตีนใหม่และโมเลกุลกรดนิวคลีอิกที่ถูกสังเคราะห์ตามโปรแกรมที่วางไว้ในโครงสร้างของโมเลกุลที่มีอยู่ก่อนหน้านี้ของกรดนิวเคลียสเดียวกัน (DNA หรือ RNA)
Biosynthesis ของโปรตีนและกรดนิวคลีอิก
การจำลอง DNA DNA เป็น Biopolymer สองที่ติดอยู่ซึ่งโมโนเมอร์เป็นนิวคลีโอไทด์ หากการสังเคราะห์จาก DNA เกิดขึ้นในหลักการถ่ายเอกสารจะมีการบิดเบือนและข้อผิดพลาดมากมายในข้อมูลทางพันธุกรรมซึ่งนำไปสู่การเสียชีวิตของสิ่งมีชีวิตใหม่ ดังนั้นกระบวนการ DNA สองเท่าจึงแตกต่างกัน วิธีการครึ่งหนึ่ง: โมเลกุล DNA ถูกทำลายและห่วงโซ่ใหม่ในหลักการของการเสริมส่วนประกอบถูกสังเคราะห์ในแต่ละโซ่ กระบวนการทำสำเนาตัวเองของโมเลกุล DNA ให้การคัดลอกข้อมูลทางพันธุกรรมที่ถูกต้องและส่งจากรุ่นสู่รุ่นนี้เรียกว่า การจำลอง (จาก lat. การจำลอง - ทำซ้ำ) อันเป็นผลมาจากการจำลองแบบสองสำเนาที่ถูกต้องอย่างแน่นอนของโมเลกุล DNA นั้นเกิดขึ้นซึ่งแต่ละฉบับจะมีสำเนาหนึ่งสำเนาของมารดา

กระบวนการจำลองแบบนั้นเป็นเรื่องยากมากเนื่องจากโปรตีนจำนวนมากเข้าร่วม บางคนกำลังปั่น DNA คู่ที่คนอื่นฉีกพันธะไฮโดรเจนระหว่างนิวคลีโอไทด์ของโซ่เสริมที่สาม (ตัวอย่างเช่น DNA Polymerase เอนไซม์) ถูกเลือกตามหลักการของการเติมเต็มนิวคลีโอไทด์ใหม่ ฯลฯ โมเลกุล DNA ทั้งสอง ก่อตัวเป็นผลมาจากการจำลองแบบแบ่งออกเป็นสองเซลล์ลูกสาวที่สร้างขึ้นใหม่
ข้อผิดพลาดในกระบวนการของการจำลองแบบเกิดขึ้นอย่างมากอย่างไรก็ตามหากเกิดขึ้นมันจะถูกกำจัดอย่างรวดเร็วโดยทั้ง DNA โพลิเมอร์และเอนไซม์ชดเชยพิเศษเนื่องจากข้อผิดพลาดใด ๆ ในลำดับนิวคลีโอไทด์สามารถนำไปสู่การเปลี่ยนแปลงที่ไม่สามารถย้อนกลับได้ในโครงสร้างและฟังก์ชั่นของ ในที่สุดโปรตีนและในที่สุดก็ส่งผลกระทบต่อความมีชีวิตของเซลล์ใหม่หรือแม้แต่บุคคล
Protein Biosynthesis ในฐานะที่เป็นนักปรัชญาที่โดดเด่นของ XIX Century F. Engels: "ชีวิตเป็นรูปแบบของการมีอยู่ของร่างกายโปรตีน" โครงสร้างและคุณสมบัติของโมเลกุลโปรตีนจะถูกกำหนดโดยโครงสร้างหลักของพวกเขา I.e. ลำดับของกรดอะมิโนที่เข้ารหัสใน DNA ไม่เพียง แต่การดำรงอยู่ของโพลีเปปไทด์เท่านั้น แต่ยังทำงานของเซลล์โดยรวมยังขึ้นอยู่กับความถูกต้องของการทำซ้ำข้อมูลนี้ แต่กระบวนการสังเคราะห์โปรตีนมีความสำคัญอย่างยิ่ง ดูเหมือนว่าจะเป็นกระบวนการสังเคราะห์ที่ซับซ้อนที่สุดในเซลล์เนื่องจากมีส่วนร่วมกับเอนไซม์สามร้อยชนิดที่แตกต่างกันและ macromolecules อื่น ๆ นอกจากนี้ยังดำเนินการด้วยความเร็วสูงซึ่งต้องมีความแม่นยำมากขึ้น
ในโปรตีน Biosynthesis จัดสรรสองขั้นตอนหลัก: การถอดความและการออกอากาศ
การถอดความ (จาก lat. transcripcio - การเขียนซ้ำ) คือการสังเคราะห์ในโมเลกุลหมึกบน DNA Matrix

เนื่องจากโมเลกุล DNA มีโซ่ต่อต้านขนานสองอันจากนั้นการอ่านข้อมูลบนโซ่ทั้งสองจะนำไปสู่การก่อตัวของ IRNK ที่แตกต่างอย่างสิ้นเชิงดังนั้นการสังเคราะห์ทางชีวภาพของพวกเขาจึงเป็นไปได้เฉพาะในหนึ่งในโซ่ที่เรียกว่าการเข้ารหัสหรือรหัสตรงกันข้าม ที่สองหรือ undogenic ให้กระบวนการเขียนซ้ำ RNA Polymerase เอนไซม์พิเศษซึ่งเลือก Nucleotides RNA ในหลักการของอุปกรณ์เสริม กระบวนการนี้สามารถดำเนินการได้ทั้งในแกนกลางและในอวัยวะที่มี DNA ของตัวเอง - ไมครอลและ Plastids
โมเลกุล Inkn สังเคราะห์ในระหว่างกระบวนการถอดความผ่านกระบวนการที่ซับซ้อนสำหรับการออกอากาศ (Iitochondrial และพลาสติก Irnas อาจยังคงอยู่ภายในอริกอยด์ซึ่งขั้นตอนที่สองของการสังเคราะห์โปรตีนเกิดขึ้น) ในกระบวนการของการเจริญเติบโตของ IRNA ที่จะเข้าร่วมโดยสามนิวคลีโอไทด์แรก (Aura) และหางของ adenyl nucleotides ความยาวของการกำหนดจำนวนสำเนาของโปรตีนที่สามารถสังเคราะห์ได้ในโมเลกุลนี้ จากนั้น STS ที่เป็นผู้ใหญ่จะออกจากเคอร์เนลผ่านรูขุมขนนิวเคลียร์
ในแบบคู่ขนานกระบวนการเปิดใช้งานของกรดอะมิโนเกิดขึ้นในไซโตพลาสซึมในระหว่างที่กรดอะมิโนติดอยู่กับ TRNA ฟรีที่สอดคล้องกัน กระบวนการนี้เป็นการเร่งด้วยเอนไซม์พิเศษ ATP ถูกใช้ไปกับมัน
ออกอากาศ (จาก lat. การแปล - การส่งสัญญาณ) คือการสังเคราะห์ของโซ่โพลีเปปไทด์บน Matrix Irnn ซึ่งการเปลี่ยนแปลงของข้อมูลทางพันธุกรรมในลำดับของกรดอะมิโนของโซ่โพลีเปปไทด์เกิดขึ้น
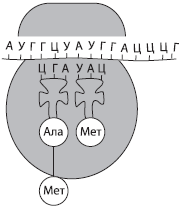
ขั้นตอนที่สองของการสังเคราะห์โปรตีนมักเกิดขึ้นในไซโตพลาสซึมเช่น eps หยาบ สำหรับการไหลของการปรากฏตัวของไรโบโซมการเปิดใช้งานของ TRNA ในระหว่างที่พวกเขาติดกรดอะมิโนที่สอดคล้องกันการปรากฏตัวของ MG2 + ไอออนเช่นเดียวกับสภาพที่เหมาะสมของสื่อ (อุณหภูมิ, ค่า pH, ความดัน ฯลฯ )
เพื่อเริ่มออกอากาศ ( การเริ่มต้น) หน่วยย่อยขนาดเล็กของไรโบโซมถูกรวมเข้ากับการสังเคราะห์ที่พร้อมของโมเลกุลของ Irnk แล้วในหลักการของการเติมเต็มไปยัง Codon แรก (AUP), TRNA ถูกเลือกถือกรดอะมิโน methionine หลังจากนั้น Subunit Ribosome ขนาดใหญ่เข้าร่วม ภายใน ribosomes ประกอบรหัส IRNN สองตัวมีให้ครั้งแรกที่ไม่ว่าง TRNA ที่สองยังถือกรดอะมิโนหลังจากที่การเชื่อมต่อเปปไทด์เกิดขึ้นระหว่างกรดอะมิโนที่ตกค้างด้วยความช่วยเหลือของเอนไซม์นั้นติดอยู่กับโค้ดใกล้เคียง ribosome ย้ายไปที่ codon หนึ่งของ irnk; ครั้งแรกของ TRNA เป็นอิสระจากกรดอะมิโนกลับมาที่ไซโตพลาสซึมสำหรับกรดอะมิโนครั้งต่อไปและส่วนของห่วงโซ่โพลีเปปไทด์ในอนาคตดูเหมือนว่าจะแขวนบน trna ที่เหลืออยู่ ไปที่ Codon ใหม่ซึ่งอยู่ใน Ribosome Trna ต่อไปนี้เข้าร่วมกระบวนการซ้ำแล้วซ้ำอีกและทีละขั้นตอนโซ่โพลีเปปไทด์จะยาวขึ้น I. มันเกิดขึ้น การยืดตัว
จุดสิ้นสุดของการสังเคราะห์โปรตีน ( การบอกเลิก) หนึ่งครั้งทันทีที่ลำดับเฉพาะของนิวคลีโอไทด์จะพบกันในโมเลกุลหมึกซึ่งไม่ได้เข้ารหัสกรดอะมิโน (Stop Codon) หลังจากนั้น Ribosome IRNK และโซ่โพลีเปปไทด์จะถูกแยกออกและโปรตีนที่สังเคราะห์ใหม่จะได้รับโครงสร้างที่เหมาะสมและถูกส่งไปยังส่วนหนึ่งของเซลล์ที่จะทำหน้าที่ของมัน
การออกอากาศเป็นกระบวนการที่เข้มข้นมากเนื่องจากพลังงานของโมเลกุล ATP เดียวถูกใช้เพื่อแนบกรดอะมิโนหนึ่งไปยัง TRNA หลายอย่างยังคงใช้เพื่อส่งเสริม Ribosomes ในโมเลกุล Irnk
เพื่อเร่งการสังเคราะห์โมเลกุลโปรตีนบางอย่างไปยังโมเลกุล irnk ซึ่งสามารถเชื่อมต่อกับไรโบโซมได้หลายอย่างซึ่งเป็นโครงสร้างเดียว - นโยบาย.
เซลล์เป็นหน่วยพันธุกรรมของการใช้ชีวิต โครโมโซมโครงสร้างของพวกเขา (รูปร่างและขนาด) และฟังก์ชั่น จำนวนโครโมโซมและคงที่สปีชีส์ของพวกเขา เซลล์ร่างกายและเพศ วงจรชีวิตของเซลล์: interfac และ mitosis Mitoz - ส่วนของเซลล์ร่างกาย meiosis ขั้นตอน mitosis และ meiosis การพัฒนาเซลล์อวัยวะเพศในพืชและสัตว์ การแบ่งเซลล์ - พื้นฐานของการเติบโตการพัฒนาและการสืบพันธุ์ของสิ่งมีชีวิต บทบาทของ Meiosis และ Mitosis
กรง - หน่วยพันธุกรรมของการใช้ชีวิต
แม้จะมีความจริงที่ว่ากรดนิวคลีอิกเป็นผู้ให้บริการทางพันธุกรรมการดำเนินการตามข้อมูลนี้เป็นไปไม่ได้นอกเซลล์ซึ่งได้รับการพิสูจน์ง่ายโดยตัวอย่างของไวรัส สิ่งมีชีวิตเหล่านี้มักจะมีเพียง DNA หรือ RNA เท่านั้นที่ไม่สามารถเล่นได้อย่างอิสระสำหรับสิ่งนี้พวกเขาต้องใช้เซลล์กรรมพันธุ์ของเซลล์ แม้แต่เจาะเซลล์โดยไม่ได้รับความช่วยเหลือจากเซลล์เองพวกเขาไม่สามารถยกเว้นการใช้กลไกการขนส่งเมมเบรนหรือเนื่องจากความเสียหายของเซลล์ ไวรัสส่วนใหญ่ไม่เสถียรพวกเขากำลังจะตายหลังจากการเข้าพักกลางแจ้งหลายชั่วโมง ดังนั้นเซลล์จึงเป็นหน่วยพันธุกรรมของการใช้ชีวิตซึ่งมีส่วนประกอบที่น้อยที่สุดในการบันทึกเปลี่ยนแปลงและดำเนินการข้อมูลทางพันธุกรรมรวมถึงการถ่ายโอนไปยังลูกหลาน
ส่วนบากของข้อมูลทางพันธุกรรมของเซลล์ยูคาริโอตนั้นมีความเข้มข้นในเคอร์เนล ลักษณะเฉพาะขององค์กรคือในทางตรงกันข้ามกับ DNA ของเซลล์ Prokaryotic โมเลกุล DNA ยูคาริโอตไม่ได้ปิดและสร้างคอมเพล็กซ์ที่ซับซ้อนด้วยโปรตีน - โครโมโซม
โครโมโซมโครงสร้างของพวกเขา (รูปร่างและขนาด) และฟังก์ชั่น
โครโมโซม (จากกรีก โครเมียม - สี, สีและ โสม - ร่างกาย) เป็นโครงสร้างของแกนเซลล์ซึ่งมียีนและดำเนินการข้อมูลทางพันธุกรรมบางอย่างเกี่ยวกับสัญญาณและคุณสมบัติของร่างกาย
บางครั้งโครโมโซมยังเป็นโมเลกุลโมเลกุล DNA prokaryotic โครโมโซมมีความสามารถในการเห็นคุณค่าในตนเองพวกเขามีบุคลิกลักษณะเชิงโครงสร้างและการทำงานและเก็บไว้ในหลายชั่วอายุคน แต่ละเซลล์มีข้อมูลทางพันธุกรรมทั้งหมดของร่างกาย แต่มันใช้ได้กับมันเท่านั้น
พื้นฐานของโครโมโซมเป็นโมเลกุล DNA สองอันที่อัดแน่นไปด้วยโปรตีน โปรตีนฮิสตั้นและโปรตีนที่ไม่ใช่ภูมิภาคมีปฏิสัมพันธ์ในยูคาริโอทกับ DNA ในขณะที่ prokaryotes หายไปโปรตีนฮิสโตน
โครโมโซมที่ดีที่สุดสามารถมองเห็นได้ภายใต้กล้องจุลทรรศน์แสงในกระบวนการของการแบ่งเซลล์เมื่อเป็นผลมาจากตราประทับพวกเขาได้รับมุมมองของลูกวัวรูปเหนียวคั่นด้วยการวาดภาพหลัก - เครื่องร้อยละ — บนไหล่. บนโครโมโซมยังสามารถเป็นได้ นกพิราบรองซึ่งในบางกรณีแยกออกจากส่วนหลักของโครโมโซมที่เรียกว่า ดาวเทียม. end Plots Chromosomes เรียกว่า telomerees. Telomers ป้องกันการทำให้เป็นเมือกของโครโมโซมและให้แน่ใจว่าสิ่งที่แนบมากับเปลือกเคอร์เนลในเซลล์พื้นฐาน ที่จุดเริ่มต้นของการแบ่งของโครโมโซมสงสัยและประกอบด้วย บริษัท ย่อยสองแห่งของโครโมโซม - chromatidผูกมัดใน centromere

รูปแบบแยกแยะความเท่าเทียมกันของโครโมโซมที่ผิดกฎหมายและเหนียว ขนาดของโครโมโซมแตกต่างกันไปอย่างมีนัยสำคัญอย่างไรก็ตามโครโมโซมเฉลี่ยมีขนาด $ 5 × $ 1.4 μm
ในบางกรณีโครโมโซมอันเป็นผลมาจากการเพิ่ม DNA หลายครั้งมีหลายร้อยและหลายพันของ Chromatide: โครโมโซมยักษ์ดังกล่าวเรียกว่า polytenna. พวกเขาพบได้ในต่อมน้ำลายของตัวอ่อน Drozophila เช่นเดียวกับในต่อมย่อยอาหารของ Ascaris
จำนวนโครโมโซมและคงที่สปีชีส์ของพวกเขา เซลล์โซมาติกและเพศ
ตามทฤษฎีเซลล์เซลล์เป็นหน่วยของโครงสร้างที่สำคัญและการพัฒนาของร่างกาย ดังนั้นหน้าที่ที่จำเป็นเช่นการใช้ชีวิตเช่นการเติบโตการสืบพันธุ์และการพัฒนาของร่างกายให้อยู่ในระดับมือถือ เซลล์ของสิ่งมีชีวิตหลายเซลล์สามารถแบ่งออกเป็น somatic และเพศ
เซลล์ร่างกาย - นี่คือเซลล์ร่างกายทั้งหมดที่เกิดจากการแบ่งมิโตติค
การศึกษาของโครโมโซมทำให้สามารถสร้างได้ว่าสำหรับเซลล์ร่างกายของร่างกายของแต่ละสายพันธุ์ชีวภาพจำนวนโครโมโซมอย่างต่อเนื่องมีลักษณะ ตัวอย่างเช่นบุคคลมี 46 ชุดของโครโมโซมของเซลล์ร่างกายที่เรียกว่า วิชชา (2N) หรือสองเท่า

เซลล์เพศ, หรือ gameti- เหล่านี้เป็นเซลล์เฉพาะที่ให้บริการสำหรับการสืบพันธุ์แบบอาศัยเพศ
Govetas มีสองเท่าของโครโมโซมมากกว่าในเซลล์ร่างกาย (ในมนุษย์ - 23) ดังนั้นจึงมีการเรียกว่าโครโมโซมของเซลล์อวัยวะเพศ haploid (n) หรือโสด การก่อตัวของเขาเกี่ยวข้องกับการแบ่งเขาออกนวลของเซลล์
ปริมาณของ DNA ของเซลล์โซมาติกแสดงเป็น 2C และเพศ - 1C สูตรทางพันธุกรรมของเซลล์ร่างกายจะถูกบันทึกเป็น 2N2C และทางเพศ - 1n1c
ในนิวเคลียสของเซลล์ร่างกายบางชนิดโครโมโซมอาจแตกต่างจากปริมาณของพวกเขาในเซลล์ร่างกาย หากนี่เป็นความแตกต่างมากกว่าหนึ่ง, สอง, สาม, ฯลฯ ชุด Haploid แล้วเซลล์ดังกล่าวเรียกว่า โพลีพอลลอย (tri-, tetra, pentaploid ตามลำดับ) ในเซลล์ดังกล่าวกระบวนการของการเผาผลาญดำเนินการตามกฎแล้วรุนแรงมาก
ปริมาณของโครโมโซมในตัวเองไม่ใช่คุณสมบัติเฉพาะสายพันธุ์เนื่องจากสิ่งมีชีวิตต่าง ๆ อาจมีโครโมโซมจำนวนเท่ากันและที่เกี่ยวข้องที่แตกต่างกัน ตัวอย่างเช่นใน Plasmodium มาลาเรียและแรงม้า Ascaris ในโครโมโซมสองลำและในมนุษย์และชิมแปนซี - 46 และ 48 ตามลำดับ
โครโมโซมของมนุษย์แบ่งออกเป็นสองกลุ่ม: Autosomes และโครโมโซมเพศ (heterochromosomes) อะทาโมส ในเซลล์ร่างกายของบุคคลที่มี 22 คู่พวกเขาเหมือนกันสำหรับผู้ชายและผู้หญิงและ โครโมโซมอวัยวะเพศ เพียงคู่เดียว แต่เป็นเธอที่กำหนดพื้นของแต่ละบุคคล โครโมโซมอวัยวะเพศมีสองประเภท - X และ Y เซลล์ของร่างกายของผู้หญิงมีโครโมโซมสองตัว X และผู้ชาย - X และ Y
karyotype - นี่คือชุดของสัญญาณของชุดโครโมโซมของร่างกาย (จำนวนโครโมโซมรูปร่างและขนาดของพวกเขา)
การบันทึกแบบดั้งเดิมของ Karyotype รวมถึงจำนวนโครโมโซมรวมโครโมโซมเพศและการเบี่ยงเบนที่เป็นไปได้ในชุดโครโมโซม ตัวอย่างเช่น Karyotype ของมนุษย์ปกติถูกบันทึกเป็น 46, XY และ Karyotype ของผู้หญิงปกติคือ 46, xx
วงจรชีวิตของเซลล์: interfac และ mitosis
เซลล์ไม่เกิดขึ้นทุกครั้งที่เกิดขึ้นอีกครั้งพวกเขาจะเกิดขึ้นจากการแบ่งของเซลล์แม่ หลังจากแยกจากเซลล์ของเด็กมันใช้เวลาพอสมควรในการสร้างอลูออนอยด์และการเข้าซื้อกิจการของโครงสร้างที่เหมาะสมซึ่งจะช่วยให้มั่นใจในการทำงานของฟังก์ชั่นเฉพาะ เซ็กเมนต์เวลานี้เรียกว่า การทำให้สุก
ช่วงเวลาจากการปรากฏตัวของเซลล์เป็นผลมาจากการแบ่งแยกไปยังการแยกหรือเรียกตายเรียกว่า เซลล์วงจรชีวิต
ในเซลล์ยูคาริโอตวงจรชีวิตแบ่งออกเป็นสองขั้นตอนหลัก: อินเตอร์เฟสและ mitosis

interfhaza - ช่วงเวลานี้ในวงจรชีวิตที่เซลล์ไม่ได้ถูกแบ่งออกและดำเนินการตามปกติ Interphase แบ่งออกเป็นสามช่วงเวลา: G 1 -, S- และ G 2-นักแสดง
g 1 ช่วงเวลา (prodittic, postmitic) เป็นช่วงเวลาของการเติบโตและการพัฒนาของเซลล์ที่การสังเคราะห์ที่ใช้งานของ RNA, โปรตีนและสารอื่น ๆ ที่จำเป็นสำหรับการสนับสนุนชีวิตเต็มรูปแบบของเซลล์ที่เพิ่งเกิดขึ้นใหม่เกิดขึ้น ในตอนท้ายของช่วงเวลานี้เซลล์อาจเริ่มเตรียมความพร้อมสำหรับ DNA สองเท่า
ใน s-party (สังเคราะห์) กระบวนการจำลอง DNA นั้นเกิดขึ้น ส่วนเดียวของโครโมโซมซึ่งไม่ได้อยู่ภายใต้การจำลองแบบเป็นศูนย์กลางดังนั้นโมเลกุล DNA ที่เกิดขึ้นไม่แตกต่างกันอย่างสมบูรณ์และยังคงถูกผูกมัดในนั้นและในตอนต้นของการแบ่งโครโมโซมมีมุมมองรูป X สูตรเซลล์พันธุกรรมหลังจาก DNA สองเท่า - 2n4c นอกจากนี้ในยุค S มีการเพิ่มเป็นสองเท่าของ Central Cell Center
g 2 ประสิทธิภาพ (postsynthetic, preasional) โดดเด่นด้วยการสังเคราะห์ที่เข้มข้นของ RNA, โปรตีนและ atps ที่จำเป็นสำหรับกระบวนการแบ่งเซลล์เช่นเดียวกับการแยก centrioles, mitochondria และ plastids จนกระทั่งสิ้นสุดการไหลบ่าโครเมียมและนิวเคลียสยังคงแยกความแตกต่างได้อย่างสมบูรณ์ความสมบูรณ์ของเปลือกนิวเคลียร์ไม่ถูกรบกวนและอวัยวะอิงที่ไม่เปลี่ยนแปลง
ส่วนหนึ่งของเซลล์เซลล์สามารถทำหน้าที่ได้ตลอดชีวิตของร่างกาย (เซลล์ประสาทของสมองของเราเซลล์หัวใจกล้ามเนื้อ) ในขณะที่คนอื่นมีเวลาสั้น ๆ หลังจากนั้นพวกเขาตาย (เซลล์เยื่อบุผิวลำไส้เซลล์ผิวหนังชั้นนอกผิวหนัง) ดังนั้นในร่างกายกระบวนการของการแบ่งเซลล์และการก่อตัวของใหม่ควรเกิดขึ้นอย่างต่อเนื่องซึ่งจะถูกแทนที่ด้วยฉัน เซลล์ที่มีความสามารถในการโทร ก้าน. ในร่างกายมนุษย์พวกเขาอยู่ในไขกระดูกสีแดงในชั้นลึกของผิวหนังชั้นนอกของผิวหนังและสถานที่อื่น ๆ การใช้เซลล์เหล่านี้คุณสามารถปลูกอวัยวะใหม่ได้รับการฟื้นฟูเช่นเดียวกับการโคลนร่างกาย โอกาสในการใช้เซลล์ต้นกำเนิดมีความชัดเจนอย่างแน่นอน แต่แง่มุมทางศีลธรรมและจริยธรรมของปัญหานี้ยังคงมีการพูดคุยกันเนื่องจากในกรณีส่วนใหญ่เซลล์ต้นกำเนิดของตัวอ่อนที่ได้มาจากผู้ที่ถูกฆ่าตายในการทำแท้งของเชื้อโรคของเชื้อโรคของมนุษย์
ระยะเวลาของการไหล่ในเซลล์ของพืชและสัตว์เป็นค่าเฉลี่ย 10-20 ชั่วโมงในขณะที่ mitosis ใช้เวลาประมาณ 1-2 ชั่วโมง
ในหลักสูตรของหน่วยงานติดต่อกันในสิ่งมีชีวิตหลายเซลล์เซลล์เด็กมีความหลากหลายมากขึ้นเนื่องจากข้อมูลถูกอ่านในนั้นด้วยจำนวนยีนที่เพิ่มขึ้น
บางเซลล์เมื่อเวลาผ่านไปหยุดการแบ่งปันและตายซึ่งอาจเกิดจากการทำงานที่สมบูรณ์ของฟังก์ชั่นบางอย่างเช่นในกรณีของเซลล์เซลล์ผิวหนังชั้นเซลล์และเซลล์เม็ดเลือดหรือมีความเสียหายต่อเซลล์เหล่านี้ปัจจัยด้านสิ่งแวดล้อมของโรค เซลล์ตายที่ตั้งโปรแกรมประหารชีวิตเรียกว่า apoptosis, ในขณะที่การตายแบบสุ่ม - เนื้อร้าย.
Mitoz - ส่วนของเซลล์ร่างกาย mitosis เฟส
ข้น - วิธีการแบ่งส่วนทางอ้อมของเซลล์ร่างกาย
ในระหว่าง mitosis เซลล์ผ่านขั้นตอนติดต่อกันเป็นผลให้ซึ่ง บริษัท ย่อยแต่ละรายได้รับชุดโครโมโซมเดียวกันเช่นเดียวกับในเซลล์แม่
mitosis แบ่งออกเป็นสี่ขั้นตอนหลัก: เคลือบ, metaphase, anafase และ telfase พยากรณ์ - ขั้นตอนที่ยาวที่สุดของ Mitosis ในกระบวนการที่การควบแน่นของโครมาตินเกิดขึ้นอันเป็นผลมาจากการมองเห็นโครโมโซมรูปตัว X ซึ่งประกอบด้วยสองโครเมติก (บริษัท ย่อยของโครโมโซม) ในเวลาเดียวกันนิวเคลียสหายไป Centrioles ที่แตกต่างจากเสาของเซลล์และแกนกลาง Achromatine (ส่วนแกนหมุน) จาก microtubules เริ่มฟอร์ม ในตอนท้ายของตรงกันข้ามเปลือกนิวเคลียร์สลายตัวบนฟองสบู่แยกต่างหาก

ใน metafhase โครโมโซมสร้างขึ้นบนเส้นศูนย์สูตรของเซลล์ด้วย centromers ซึ่ง microtubules ติดอยู่กับการแยกกระดูกสันหลังที่เกิดขึ้นอย่างสมบูรณ์ ในขั้นตอนของการแบ่งโครโมโซมนี้พวกเขาถูกปิดผนึกมากที่สุดและมีรูปแบบลักษณะซึ่งช่วยให้คุณศึกษา Karyotype
ใน anafhase การจำลองแบบอย่างรวดเร็วของ DNA เกิดขึ้นใน Centromers ซึ่งเป็นผลมาจากการแยกโครโมโซมและ Chromatids ไม่เห็นด้วยกับเสาของเซลล์ที่ยืดออกโดย microtubes การกระจายของ Chromatide จะต้องเท่ากันเนื่องจากกระบวนการนี้ทำให้มั่นใจได้ว่ามีการเก็บรักษาความเป็นมาตรฐานของโครโมโซมในเซลล์ของร่างกาย
บนเวที สายพันธุ์ โครโมโซมเด็กจะถูกเก็บรวบรวมบนเสา, disirates เปลือกนิวเคลียร์จากฟองอากาศรอบ ๆ พวกเขาและนิวเคลียสปรากฏในเมล็ดที่เพิ่งก่อตัวใหม่
หลังจากหารนิวเคลียสไซโตพลาสซึมถูกแบ่งออก - ไซโตไคน์ ในระหว่างที่มีการกระจายตัวมากขึ้นหรือน้อยลงของอวัยวะทั้งหมดของเซลล์แม่
ดังนั้นอันเป็นผลมาจากการ mitosis บริษัท ย่อยสองแห่งจะเกิดจากเซลล์แม่หนึ่งตัวซึ่งแต่ละแห่งเป็นสำเนาทางพันธุกรรมของมารดา (2n2c)
ในผู้ป่วยที่เสียหายเซลล์อายุและเนื้อเยื่อเฉพาะของร่างกายส่วนที่ค่อนข้างแตกต่างกันสามารถเกิดขึ้นได้ amitosis พวกเขาเรียกกองโดยตรงของเซลล์ยูคาริโอตซึ่งเซลล์ที่เทียบเท่าพันธุกรรมจะไม่เกิดขึ้นเนื่องจากส่วนประกอบของเซลล์จะถูกแยกออกจากกันอย่างไม่สม่ำเสมอ มันเกิดขึ้นในพืชใน Endosperm และในสัตว์ - ในตับกระดูกอ่อนและกระจกตาของดวงตา
meiosis ขั้นตอน Maizia
meiosis - นี่คือวิธีการของการแบ่งทางอ้อมของเซลล์อวัยวะเพศหลัก (2n2c) ซึ่งเป็นผลมาจากเซลล์ Haploid (1N1C) ที่เกิดขึ้นส่วนใหญ่มักจะมีเพศสัมพันธ์
ซึ่งแตกต่างจาก Mitosis, Meyosis ประกอบด้วยสองหน่วยงานต่อเนื่องกันซึ่งแต่ละแห่งจะถูกนำหน้าโดย Interfac ส่วนแรกของ Meiosis (Meios I) เรียกว่า การลดลงเพราะในขณะเดียวกันปริมาณโครโมโซมเพิ่มขึ้นเป็นสองเท่าและส่วนที่สอง (MEIZ II) คือ สมทบตั้งแต่ในกระบวนการจำนวนโครโมโซมจะถูกเก็บรักษาไว้

INTERFAZ I. รายได้เช่น mitosis interfase Meiosis I. มันแบ่งออกเป็นสี่ขั้นตอน: Poarase I, Metaphase I, Yanatherapy I และ Bulpase I ใน profare I. กระบวนการที่สำคัญที่สุดสองกระบวนการเกิดขึ้น - การผันคำกริยาและ crossfield การผันคำกริยา - นี่คือกระบวนการของการรวมโครโมโซมที่คล้ายคลึงกัน (จับคู่) ตลอดระยะเวลาทั้งหมด คู่โครโมโซมที่เกิดขึ้นระหว่างการผันคำกริยาจะถูกเก็บรักษาไว้ในตอนท้ายของ Metafase I.
cross hopeer - การแลกเปลี่ยนร่วมกันของส่วนที่คล้ายคลึงกันของโครโมโซมที่คล้ายคลึงกัน อันเป็นผลมาจากกากโครโมโซมที่ได้รับจากร่างกายจากผู้ปกครองทั้งสองได้รับการผสมผสานระหว่างยีนใหม่ซึ่งทำให้เกิดการเกิดขึ้นของลูกหลานที่มีความหลากหลายทางพันธุกรรม ในตอนท้ายของ Profase I เช่นเดียวกับใน Mitosis Profhae นิวคลีโอลัสหายไปแกนหมุนหายไปที่เสาของเซลล์และเปลือกนิวเคลียร์สลายตัว

ใน metafhase I. คู่โครโมโซมถูกสร้างขึ้นบนเส้นศูนย์สูตรของเซลล์ microtubule ถูกแนบกับ centromeres ของพวกเขา
ใน anafase I. โครโมโซมที่คล้ายคลึงกันทั้งหมดประกอบด้วย Chromatids สองตัวที่แตกต่างจากเสา
ใน Bulfase I. รอบกลุ่มของโครโมโซมในเสาเซลล์ที่เกิดขึ้นเปลือกนิวเคลียร์นิวเคลียสจะเกิดขึ้น
Cytelnez I. ให้การแยกพลาสซึมของเซลล์เด็ก
เซลล์ย่อย (1N2C) ก่อตัวขึ้นเนื่องจาก Meiosis เป็นความแตกต่างทางพันธุกรรมเนื่องจากโครโมโซมของพวกเขาบันทึกแบบสุ่มไปยังเสาเซลล์มียีนที่ไม่เท่ากัน
ลักษณะเปรียบเทียบของ mitosis และ meiosis
| เข้าสู่ระบบ | ข้น | meiosis | |
| เซลล์ใดที่เข้าสู่การแบ่ง? | Somatic (2N) | เซลล์เพศหลัก (2N) | |
| จำนวนแผนก | 1 | 2 | |
| มีกี่เซลล์ที่เกิดขึ้นในระหว่างการแบ่ง? | 2 somatic (2n) | 4 ชั้น (n) | |
| interfhaza | การเตรียมการของเซลล์สำหรับการแบ่ง DNA เป็นสองเท่า | สั้นมาก DNA สองเท่าไม่เกิดขึ้น | |
| เฟส | Meiosis I. | {!LANG-2db15907e81683fbe68b6c6787718d01!} | |
| พยากรณ์ | {!LANG-cf2bc9b2d230d3dc5264c1473bc50db7!} | {!LANG-c909ae37af80d118b6cc4daa78a7ba01!} | |
| {!LANG-b3bf330b836d4268ad7d369831cf9da1!} | {!LANG-ea4632ccbe8a2281477bbad81697252f!} | {!LANG-5daf3fe2720ee2205d6b4b79d5a8375b!} | |
| {!LANG-c84a97d11b6001fdbccbfddbdea9681f!} | {!LANG-e4122f5b2de632b444f25cf3b7718bad!} | {!LANG-8252082aa8bb158944155e43a3b23678!} | |
| {!LANG-5960bc79c71567903232e45cf3628a90!} | {!LANG-bd51064b943d366740e6af22e975f111!} | {!LANG-bd51064b943d366740e6af22e975f111!} | |
{!LANG-6b744301453cea5469a64b486b5db894!}{!LANG-346828220b028a3bc9f78dc5525a4ca4!}
{!LANG-2db15907e81683fbe68b6c6787718d01!}{!LANG-0a05689e2c3d67237892e81a28a77592!} {!LANG-e94ed134a0dd93843dfb4628c104aad6!}{!LANG-b074771cf4f2cb847365e028f64fb06b!}
ใน {!LANG-3a12a92973d1b40ab436a51d372cb1a3!}{!LANG-0840acbaf23dee482a19b04482303349!}
ใน {!LANG-f63063a7a56387c20563125468b61abe!}{!LANG-99077673f49827a201e25bd73cefb5d2!}
ใน {!LANG-0551284f9845ba95336579b3bbda81b1!}{!LANG-b446b6b67093fd7b92b0421edfab57d5!}
{!LANG-3c1d0cb0a3bd2d5ce871eb4a13d2c3da!} {!LANG-83029384967a6f0a770e687bec9ff52b!}{!LANG-929e4ddd757eaeaa21b6b927f08137a4!}
{!LANG-135737a53027db1ee2fc3ea88c2402ed!}
{!LANG-c621d76ba7e4a4e0f234ddb21b7ee633!} (จากกรีก {!LANG-1886c31da564441ca959866b5d546817!}{!LANG-c77987e7deb6a9c2319aa6a6de993275!} {!LANG-b9337ebc8378942ba7c95dd56295ddbe!}{!LANG-58758dc5f41e67ce5817c112506ec6f7!} {!LANG-1ce0cc26668c040dd54621ed79a17e69!}{!LANG-54a22154b92775a8101a187321673438!}
{!LANG-53f81acf8b34a13902fab82f66604909!}
{!LANG-799f3bdbd5dfdda206fe6bc1032d2672!}
{!LANG-af1fea84be25cba68a8fa9d4f1b7d43d!}{!LANG-ef4111175d9d85db3fb2dbd066aad284!}
{!LANG-98e5a471b855c5212e1d71b08c79254b!} {!LANG-dc7ebe125dfcea06a449dc2079615dcd!} (จากกรีก {!LANG-f234b9849d18daf068bacee029ef068c!}{!LANG-a111f73f42190e1eee3ccff68e50a327!} {!LANG-c0724e019984932867bde34bfb3f9498!}{!LANG-4eb3dbedd91327416d86482e04b0905b!} {!LANG-36d57753071000bd46bf16e6c1163701!}{!LANG-19e6c63a233c070ea117d5a4d3814644!} {!LANG-6ad4afb08b45766d447efd06661caed5!}{!LANG-e8dee1df43ad0356ed41c4a083aba9b5!} {!LANG-1bf0761e6f1330e1bbf1d7cc8a4a5b92!}{!LANG-3f26b292d6acdf356aee10f500f8225d!}

{!LANG-b142fc99732740572cb91e24253af982!} {!LANG-3a1d1ec0368a07123a0657f545a16c05!} (จากกรีก {!LANG-cb88061e0fa04c8ebfe91af63020e2b0!}{!LANG-84f21a25ab83a3cfe87202b7a0dd175a!} {!LANG-e2b2c50bb0ada72a51b0d48162978814!}{!LANG-0fca98af138d38fac0950d9bb5ec0b46!}
{!LANG-bcf68256ef34a3fdcd835083ca3f78a1!}{!LANG-c92cd50312b12a2bd20b82fc10a18317!}
{!LANG-2f82e299f7fc7c6494119437f2bf0193!} (จากกรีก {!LANG-b0ab1ba45bf45614f5d8958e0c26197e!}{!LANG-d86c9c20c66df6dd839ec8fbcdb8c386!} {!LANG-1ce0cc26668c040dd54621ed79a17e69!}{!LANG-e06c86fd3f15004415fb68cf14a70464!}

ใน {!LANG-82b3a50bd4e7fbc053317aebbcfb7327!}{!LANG-9f8c7c0328463976e0649f5315d69f85!} {!LANG-62d8629de62d9b5f8560f970fdfc6c80!}{!LANG-b66e7912f218a64cc4f3427f85b7090d!} {!LANG-98c4d9075a198c2d318eef4f3028e795!}{!LANG-11b5aa3d5ae18e3486957cb0d164baa2!} {!LANG-777af8b8fe6d4a4d34320021cc51d0cb!}, หรือ {!LANG-52aea3d47b7e0cc6864f4939a0c0fe43!}{!LANG-db2ac7cec9b75c5bef245e6f5b1efefc!} {!LANG-9ec04b6c5bf7406172908608ca7d2d41!}{!LANG-0ab0131e79aa66d5f388721402166918!} {!LANG-a96a897b68b33b039629977478ce84c4!}, หรือ {!LANG-ab9b455bd41f6cc916db922600d747b7!}{!LANG-4dd585122201ef8bca7742ee82763254!} {!LANG-d6c33d9b5ad702fce35af4a03c50ce12!}{!LANG-b66e7912f218a64cc4f3427f85b7090d!} {!LANG-dc9f5894450e03eeec4ae58a88abcd8b!}{!LANG-78bcd27226af2c64b592e0421259466c!}
{!LANG-2a31bd44eb964278719786d75571aa62!}, หรือ {!LANG-eecce7626afe0146489b18f315ad19ab!}{!LANG-0e2e0377613ab283e05c6e2babd377bc!}

ใน {!LANG-d2c377337bb1a7c56434d94562c25495!}{!LANG-d06e3db809b7f2a269e931644fa83744!} {!LANG-f9cfaf6d33a40b39cf55b0c947cbd4d8!}{!LANG-9acfcfc6e75ebb504dd6180c5749d916!} {!LANG-f43f7851a2a4ac59f43fa619f2224c53!}{!LANG-92c1baa162d16adb8cd7635f5176487a!} {!LANG-e4dff48600b78e55e1967750dd546299!}{!LANG-311c312faec965c63127423256161ca7!}
{!LANG-d210eba5cd02881ef183ab48a19c8d2d!} (จากกรีก {!LANG-e3c53f5cb30e98994331103b1458c58c!}{!LANG-24abad78a79283e1a8887a81a1db8e9a!} {!LANG-1ce0cc26668c040dd54621ed79a17e69!}{!LANG-72ffcc4932a1a4d0fc2afcb0904f9b2a!} {!LANG-3579e4b89b089f78ed97236ec2094cbe!}{!LANG-f08738031e7bdcd16fb6bd30724ba922!} {!LANG-8dca97af7867a4c467ed8ea96fa0c6a9!}, หรือ {!LANG-99e85e8ce9c8f10979269b9a292a0e46!}{!LANG-6031606f291cf7ccbcbbc26b2db0d60d!} {!LANG-9ec04b6c5bf7406172908608ca7d2d41!}{!LANG-e9d573c603158a82903396100c7cff02!} {!LANG-3aedc6643ab1a00abc09e522f239a0b2!}, หรือ {!LANG-5b86880d2a26c0975fda5a8555e3433f!}, ผม. {!LANG-e5765b17dbaa97825c12e99337861b67!}{!LANG-2a139e117edbd82593e9b8271315b36e!}
{!LANG-a225800dbd90ae80e2583978af42d2b1!}
{!LANG-13ba75689299bbb1278bc1adf414103d!}{!LANG-e5e275ab701740fca7eff4767a70a39c!}

{!LANG-4d4f60ac330735f3d42fbe90064533e3!}
{!LANG-31d47247dd1b1a5e2fcb8566762f5693!}
{!LANG-c1390f51f8ff1f938052749dadf43b49!}
{!LANG-cd022f6dfc908abc63383ebfbb298703!}
{!LANG-81b7ebcd542f0e48b410ae5a9eda0d39!}
{!LANG-846b1a57a7b44e72881123e95d6a55ac!}
{!LANG-6c8551c8df802de3a38e9c72b5d0765d!}
{!LANG-38589e5dc7a00d00adb86307bbc85b98!}
 {!LANG-3eeb9ace90b155651273a991ae02835c!}
{!LANG-3eeb9ace90b155651273a991ae02835c!}
{!LANG-ac1f127d4e0024cd560efa9c655f670c!}
{!LANG-395a33ba265dbf1fa16274e8639a7c55!}
{!LANG-8f0600010944537ee4298e440aae88bf!}
{!LANG-620c59eaa6a5c1fd1efa8ffb350a0449!}
{!LANG-bd134194d4b40ac0330bd9371b4269b4!}
{!LANG-f2fccbf8e0f86eda42181048bb551c38!}
{!LANG-31496e69e883b2800b5e547076bd2107!}
- {!LANG-6c3b47bfacf9a694a6f268385bb70791!}
- {!LANG-22917b592e4807989140134f67e2a5bc!}
- {!LANG-f5bec47d88f44d25b645464a45db7be6!}
- {!LANG-132fa436f220aa6a835b00539920315f!}
{!LANG-d86ea87d8a13b524890eb28f3a2565c8!}
{!LANG-a2e9ea81a160c6dea669ca29ced5466a!}  {!LANG-43154bd8609eb3a75008700d5a138b45!}
{!LANG-43154bd8609eb3a75008700d5a138b45!}
{!LANG-54449569c48aed59cc6c5240e01a3e63!}
{!LANG-31496e69e883b2800b5e547076bd2107!}
- {!LANG-4108774502967cdc6855939cc1decfc1!}
- {!LANG-e31910923851da4b807a09b00c6bd9ed!}
- {!LANG-96b068bc0a18c144b826d0ff8c77dd9e!}
{!LANG-ce475d7ea40ba2f00ee2ad580267c3d5!}
{!LANG-82f883b77c361eb3e750e5013818815c!}
{!LANG-c613955a0e3b35df946fe63301600923!}
{!LANG-31496e69e883b2800b5e547076bd2107!}
- {!LANG-009113c7582053d3bcccceea373cb071!}
- {!LANG-47aa34e6ce8c6dd06ca15202d25c7cd0!}
- {!LANG-d1f9471ea3b717c03eb6f6018dd8a059!}
{!LANG-00ed989bf103b82c0daa9125dcec8b1e!}
{!LANG-65c8cd05562ed458affd063d9e79d1cc!}
{!LANG-fae31f8facf204214bcd5651260626ef!}
{!LANG-31496e69e883b2800b5e547076bd2107!}
- {!LANG-4c8145fc2989dd3255b933a480550f52!}
- {!LANG-ebf088ad6ff0526b0deaaecc5f0f5098!}
- {!LANG-6b7a24caba5ecda2696dc66d1b830b80!}
{!LANG-31129f975d59e17e8395c139f24700f5!}
{!LANG-d6714e5720b1387a978d43737dfbcb65!}
{!LANG-64e8e0d0d0868d06018d9f1bb1214e1a!}
{!LANG-31496e69e883b2800b5e547076bd2107!}
- {!LANG-95ed4783c3a369d666d3c8f0f1df398a!}
- {!LANG-c97b595d27ae00b16c67142794c4f16b!}
- {!LANG-da26137e871a5fb47c75f683097c2e38!}
{!LANG-d6f233c017903e19995964f7016069aa!}
{!LANG-ec7c61a028b485f83fa7fdcf7b9e1b85!}
{!LANG-fba4f2e29865a4fec397548c7087be98!}
{!LANG-31496e69e883b2800b5e547076bd2107!}
- {!LANG-f33f091eb6298800a867e121159a34f7!}
- {!LANG-fc5e00800ceacb5ad1648da0d4a17038!}
{!LANG-a29d9f2fffb47a4ca7f043960a94fc2c!}
{!LANG-b1c3e9165f0bdeb62d9ca922bd8e8f8d!}
{!LANG-45208e856dfbf9c662d5f26cc6482e7a!}
{!LANG-31496e69e883b2800b5e547076bd2107!}
- {!LANG-74f6f22d30087596ff079f530696329e!}
- {!LANG-d23b8c5a89e90722dabd728ea42b204d!}
- {!LANG-9e99e6ea2ddacac0210e2726e53f753f!} {!LANG-1bf3b9caa6690a6091434415c871391e!}{!LANG-d9b58dd8bb78c284d5cc8f43b20f6257!}
{!LANG-cb73491864e0c209b3abe5e1a7d73b44!}
{!LANG-66462094c3d00b3ccfedc7b7a3a4ce3b!}
{!LANG-e4a4b4f18259654b3fbef5aad5db5362!}
- {!LANG-6ed75f097d0f31c09c900fa53ba0d813!}
- กรดอะมิโน, แฝดสามนิวคลีโอไทด์
- Triplet กรดอะมิโนโมเลกุล T-RNA
- {!LANG-c079506114d9b6444ab6aa9b382b4136!}
- ชิ้นส่วน DNA: Tsgattsaagahatg Anticodones T-RNA: CGA, Uua, CAA, GAA, ส.ค. ลำดับกรดอะมิโน: Ala-Asn Val-Lei-tir
- {!LANG-ca6a303fd27936bb588f243ce57c7865!}
- . ชุดพันธุกรรม:
- {!LANG-c30ea7e728f282c3b1ac28fdd9612be2!}
- {!LANG-24546e44865018a5b1a3bf593cf5e3b0!}
{!LANG-613d95e7e762bc66058ab1bbf860073c!}
{!LANG-ff83018f65bdcd84cc585345b8e03c3a!}
{!LANG-de66749a65c6eb9e152d3568dace1368!} {!LANG-caa7732b265874d264bdd9622a90b916!}{!LANG-2596c70d7e7ceac2f2dbdbe2deed8a0a!}
{!LANG-58af8a3751fc05ab79dc09a162945edc!}
{!LANG-5118b27112adbf107e76889a51ea581c!}
{!LANG-7f24902b4269900accdc6692b9d25deb!}
{!LANG-7f0986e1d047280000e47e5a8c00db4a!}
{!LANG-50f926a5ebdbe262cfaa6f4518f0d732!} {!LANG-8198c6c9e8c6e3ce455fbc6f24104d37!}{!LANG-0e3b411898e7e9728393d22b71378aa4!}
{!LANG-0d365b9c2199bae5d62476f4b2e282db!}
{!LANG-ed0a553d01b4a4a72ce0af8a47dbe119!}
{!LANG-c38fcc5d275dffb06fd8904dd9e54e31!}
{!LANG-df3c6f357d9c38fade480e9a4deccd35!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-e9d1364e16e2fd2d9ba9f325e0e19f02!}
{!LANG-04421b0191445f9720d9f04feb449eea!}
{!LANG-f010d8a77512361fa5fca060d56e97da!}
{!LANG-2911bef6697c0dc48f04e2ee0b00ad05!}
{!LANG-5db1bed965afb40457af9e699bba2cb7!}
{!LANG-a6041f3cf55a4f4648e00434f4c0922b!}
{!LANG-0649d3dbb6168b8b681abb7ecc010be6!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-43228e9f842ac675b6734ea1c5854fe0!}
{!LANG-998144b86ff38f217bd8e6cc478a9ed4!}
{!LANG-a125a580771215e245f2005eea6d90a8!}
{!LANG-88e30626f91d4e5940a7fc32b2f85fee!}
{!LANG-8eaa441d8eb3bbc0255db086d8e8d654!}
{!LANG-f3494279b76c2041b4838534778db035!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-64a00420946da0241d0de6c1bcb3fabd!}
{!LANG-f55686778fefebe5d0cf3481313a0b11!}
{!LANG-4d6e43fc5a60920196e6ea24e05c08e0!}
{!LANG-a125a580771215e245f2005eea6d90a8!}
{!LANG-47189a30c49c3aa9024631b527eeddfe!}
{!LANG-cae0ff5d22d8a8dbc906d3db0f3fb54f!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-54f40ae2f7fb1fe68abc56939a434742!}
{!LANG-a9a880c18d74bdcd0a930fcc976a40f0!}
{!LANG-3b5217781af4514a6159e1bcaa1807ef!}
{!LANG-a125a580771215e245f2005eea6d90a8!}
{!LANG-0361b2529f99d598bb6216f3a82cb177!}
{!LANG-ce4fa2c19588c662f2eee5917bd9ac86!}
{!LANG-231ef15f3356c9c3fa5525c577fa0cca!}
{!LANG-3f18996b5cef5dcbadfa70e85c34f97f!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-3a830fcb190e39c5d4f4b772e3ca8b7e!}
{!LANG-e2c05013efd456811b7f73de2103aa89!}
{!LANG-a125a580771215e245f2005eea6d90a8!}
{!LANG-0de9d7a25e663d779899502273296e3c!}
{!LANG-4f624a34cb42bc722a2c7698e675eeea!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-91b723dc5277557dc12e162faa8de60e!}
{!LANG-29aa604b5a26392aecefa66aa62bd9a8!}
{!LANG-9ed7d00e49a72306a6124d97ba2f47c6!}
{!LANG-a125a580771215e245f2005eea6d90a8!}
{!LANG-3f070429507764dddd411e8f5cf8db4b!}
{!LANG-52e4a7aac5a7424f758cecbb34fcb105!}
{!LANG-4dcdc3c209a14abde4c1cf2c2fdf4115!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-36792a53f4d2780116532293978dd6d4!}
{!LANG-d39208dfda28beca89065e762d854369!}
{!LANG-a125a580771215e245f2005eea6d90a8!}
{!LANG-78861c62105ef4fcb84bdc67f2ba93a9!}
{!LANG-ffc739c3898c440f1c01951be1622be9!}
{!LANG-739ac9341c7f1ecc0232c7e66f9440e2!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-140aa78ddb66281aedd5f2ead331c1e1!}
{!LANG-08e9d17ed00604fe3f51b2075eb7d4b0!}
{!LANG-a92acd99160611e6ae3772a83d8a9a19!}
{!LANG-a125a580771215e245f2005eea6d90a8!}
{!LANG-ab3f611fb99fa4e3dc30fd6d118ed15e!}
{!LANG-1da8eb272c81c60a59db2e1fe7fa2bfd!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-6596c53fe4b2b835aaa08055897dc95e!}
{!LANG-0a92f7817992393ea984c8a539e3b020!}
{!LANG-6fc352b3cc31fe3357df0ebac6607061!}
{!LANG-a125a580771215e245f2005eea6d90a8!}
{!LANG-d8ca79459547f231c54650222fb851da!}
{!LANG-4375edb79dda144ae17bb9aa76b50618!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-58ceb98ff1a21e3fb4e34c8888e1d5d8!}
{!LANG-1ad183f85ad56c6de5fc77651423bd27!}
{!LANG-a125a580771215e245f2005eea6d90a8!}
{!LANG-dae223912c6e72e15a3fe3386bc22b19!}
{!LANG-1b6993f2352791649d3fb53848f4c06e!}
{!LANG-113736576fe21d7f014c14e070bc9f5d!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-c6ceeb2f95a6cacc31d2531504704da2!}
{!LANG-1aae8d949243d201677c77503adccfa2!}
{!LANG-4bd9850721724736fc8b179574c79d7a!}
{!LANG-a125a580771215e245f2005eea6d90a8!}
{!LANG-bdc576e3533f79d9f8832b17d4ca6b5d!}
{!LANG-6059bdca022d5e226a70369ab785582c!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-afcea80faf07673c0c6dca5ea32cb33a!}
{!LANG-0b1eadcf811b60ead23bdeb4e29fb6f6!}
{!LANG-a4937f7ed4521a811da703847ca18bcd!}
{!LANG-c7563e9a316111ce69f5d2615534dca4!}
{!LANG-a125a580771215e245f2005eea6d90a8!}
{!LANG-b97dcdc984c53cccca3f9639bbd4bf27!}
{!LANG-0b95c7ea92c95d2deb06dd077d68f455!}
{!LANG-ed1cf3dba613b0302ee27ca650a0a7bb!}
{!LANG-a412c7e071dbbbb07fcf6855365e0c4c!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-402b4c42af8d7da91f30edd962cc3a67!}
{!LANG-bc48b222fa79bee2e3bf6dd58cc2ab6b!}
{!LANG-7e82074f75e908d43f56760c448ed590!}
{!LANG-237edd35a56e3e9bc2be612dff020d8d!}
{!LANG-a125a580771215e245f2005eea6d90a8!}
{!LANG-1932f0c7af5cbb3a9b8d7e72d9e8204f!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-415f8a8222d36dd9b701028072e31ac3!}
{!LANG-b960aee03713854b863c0f23f14a6002!}
{!LANG-afd2443a6beaf19b9d9cf760422f2e71!}
{!LANG-a125a580771215e245f2005eea6d90a8!}
{!LANG-4b2b18ba7af87c844ae68528c678de25!}
{!LANG-16efe89080da3fa2a828ccb08d8a685b!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-dd39abb0a32836895d0b47f98af9ce9d!}
{!LANG-fa27db975600265123d261f0d19e0cd9!}
{!LANG-a125a580771215e245f2005eea6d90a8!}
{!LANG-18cb128ccbaa18fd6dc6157c5ca3077c!}
{!LANG-61a6a1e0130fd3771b075a1137f2d481!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-b24f6d5370298ff6487424f349b7bb58!}
{!LANG-13ccf9269fa04cbb5d51c9d5af093a74!}
{!LANG-a125a580771215e245f2005eea6d90a8!}
{!LANG-2d2f0359ea84d835677471fc1face0f5!}
{!LANG-345dad76931a4e2d75813e894f1b3e61!}
17. {!LANG-4eeca70648793b15bd388fafb2dd4528!} |
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-702b052177308b95d40498fd2ca1ca7b!}
{!LANG-8aea766e55e34bf22ea5c658d5b03cc8!}
{!LANG-a125a580771215e245f2005eea6d90a8!}
{!LANG-39c36a39cbe11442fca34dfa2415bdd5!}
{!LANG-afcd9f2c4cbc11c0c49ff94ee08d2b16!}
{!LANG-3bf64ef155a174438b0bc6523662d11a!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-5b70a8c89bad8b0443466da1d5f83e06!}
{!LANG-d8b6fcd3917e0b27cfa798d571511e1b!}
{!LANG-4ba5254873dad1d4ad11e7d977d40ce0!}
{!LANG-a125a580771215e245f2005eea6d90a8!}
{!LANG-1838dd0c71297694a9164df294ecb5ba!}
{!LANG-22b7705c6e5523cec6f7a9c85ea1af60!}
{!LANG-33b1b9f80a92a71b57c27443ade26741!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-8716d1dcfa87334199920e8bef3baf1b!}
{!LANG-436b093145272f1298bcd29cbcdcf88d!}
{!LANG-a125a580771215e245f2005eea6d90a8!}
{!LANG-2db6601eb39c88fb368e925acecb9fab!}
{!LANG-9e66732903d3d1c1984f3e66350f9b73!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-ca81001de3ac6dfc8ff42821a470475c!}
{!LANG-adc2367b7fc884b9d3c63c8099eb21c8!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-ea010c9f1d6d611d56b6a21e8a29be22!}
{!LANG-c3092aa02e0618029a282bbd263947fd!}
{!LANG-a125a580771215e245f2005eea6d90a8!}
{!LANG-07cce984bdb977623cb76d0b7c45bdd4!}
{!LANG-f323a0fdbe7c324e4316c3750262aa83!}
{!LANG-fd694502f1fd6c984f07df3a2c0c596b!}
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-091dd1718e989b9e8e4ed259cc9a0626!}
{!LANG-7043a078e16762e2dea64afd4c0df466!}
{!LANG-45022d621011833937cc8905f20d1bd2!}
{!LANG-9cbaf03a88931ce3b187dd38b07cc20d!}
2. {!LANG-5d88e141ceeebc7008fd83e79e7ace93!} {!LANG-68f2e9e4eace608887f4608eada5179b!} |
{!LANG-b20b9fe939d805c442802b4a6f9d20a2!}
{!LANG-63deb6078426d42e93d1f282074bebc8!}
{!LANG-b1852a6b96c4dc1baecdbe13dc151216!}
{!LANG-3e3ca3c28ff47cb4a122b70dff042402!}
{!LANG-8afb68df35abb4cb289db74669b0251e!}



